Презентация "Основы генетической инженерии" – проект, доклад
 Слайд 1
Слайд 1 Слайд 2
Слайд 2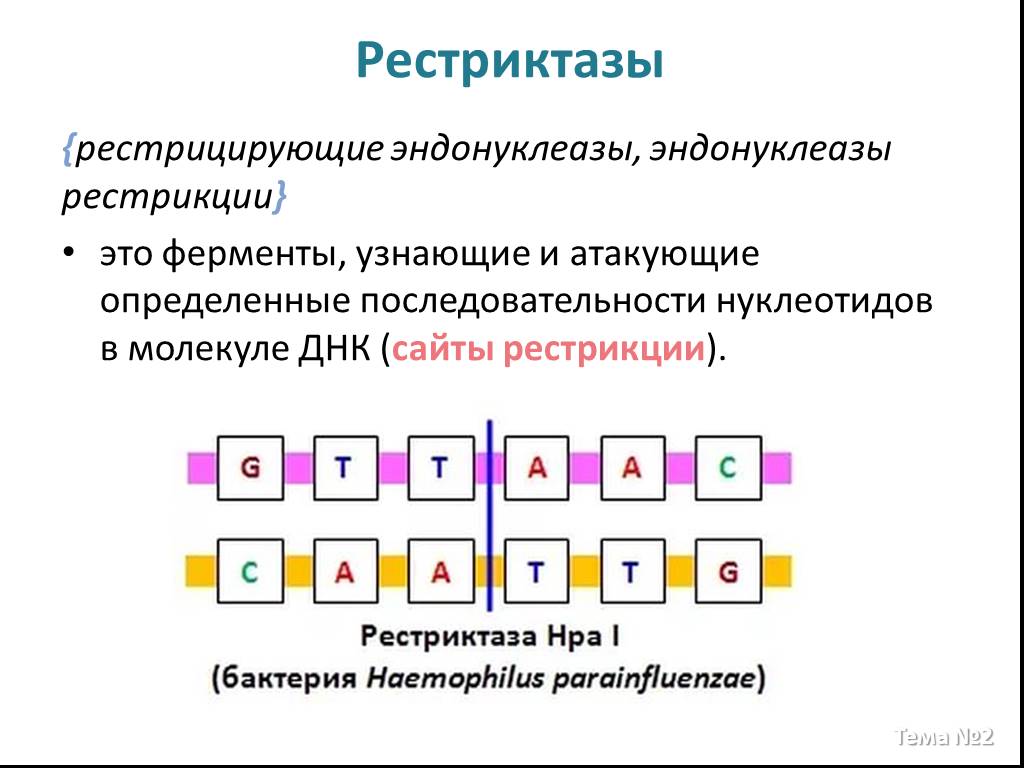 Слайд 3
Слайд 3 Слайд 4
Слайд 4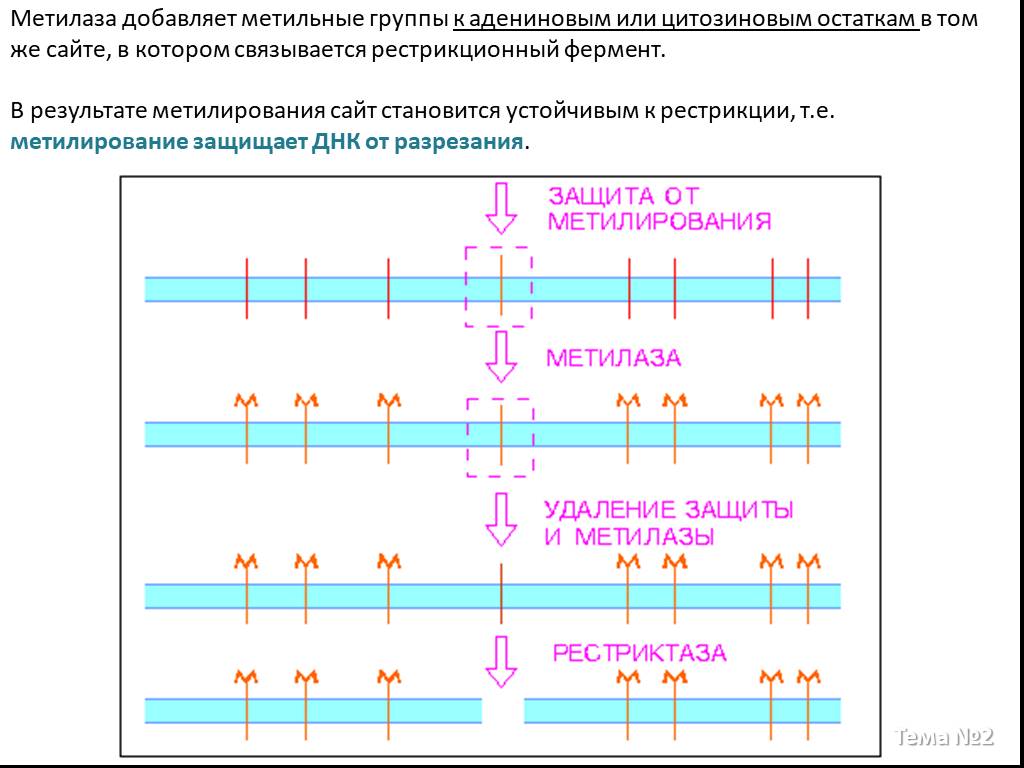 Слайд 5
Слайд 5 Слайд 6
Слайд 6 Слайд 7
Слайд 7 Слайд 8
Слайд 8 Слайд 9
Слайд 9 Слайд 10
Слайд 10 Слайд 11
Слайд 11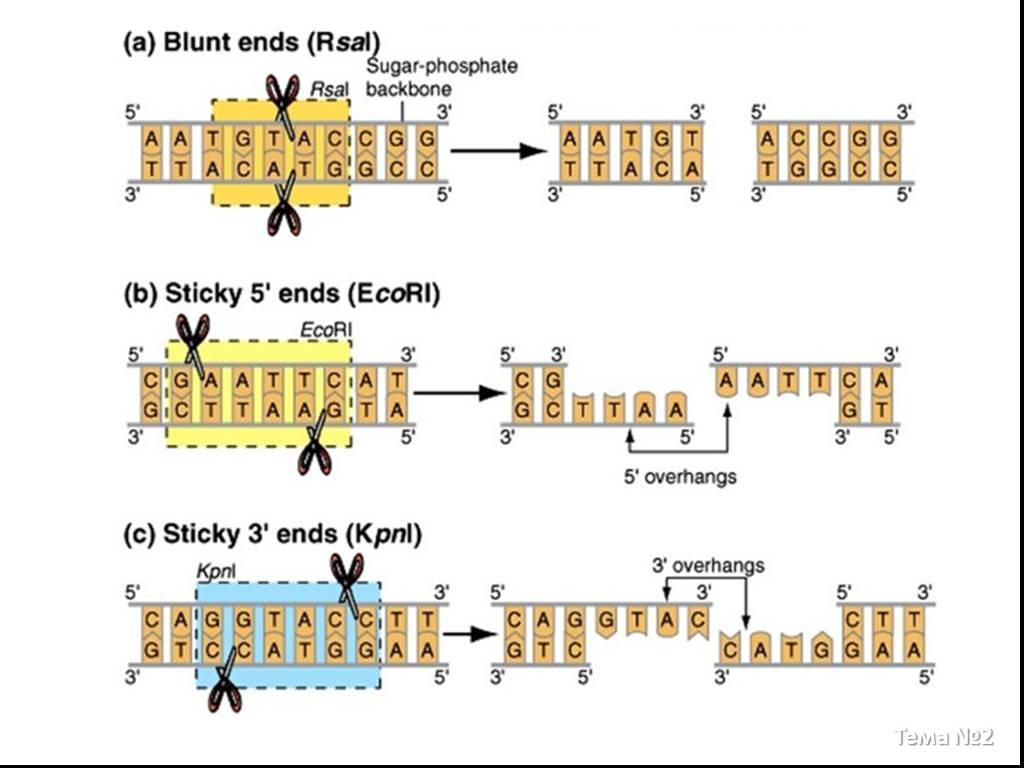 Слайд 12
Слайд 12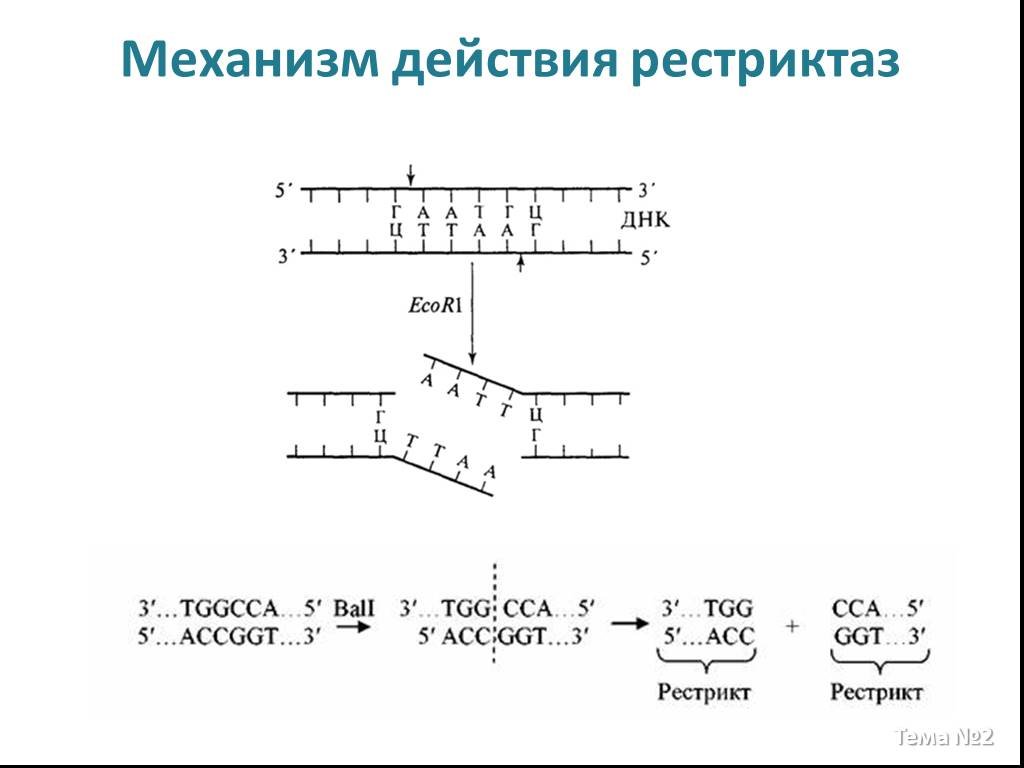 Слайд 13
Слайд 13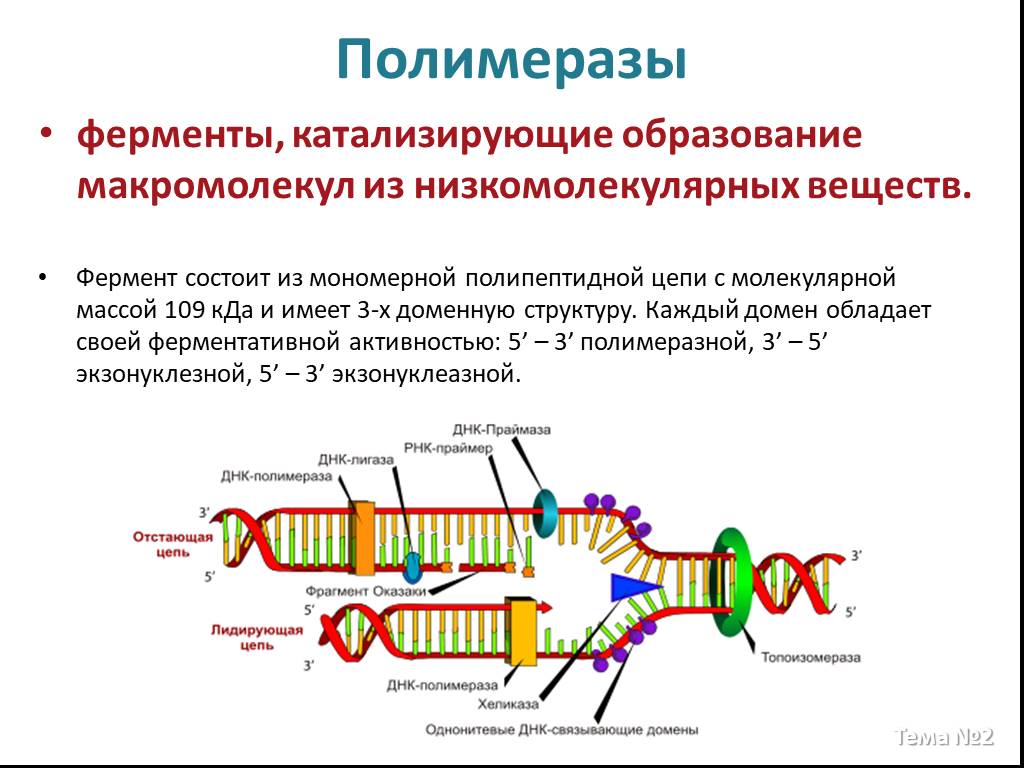 Слайд 14
Слайд 14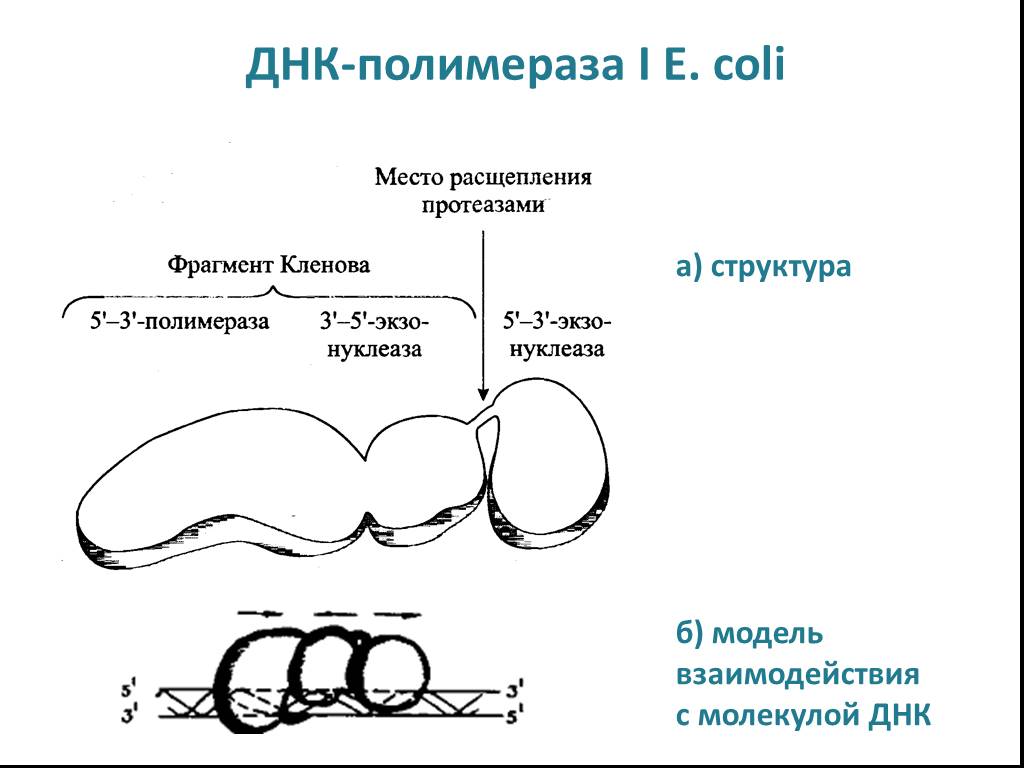 Слайд 15
Слайд 15 Слайд 16
Слайд 16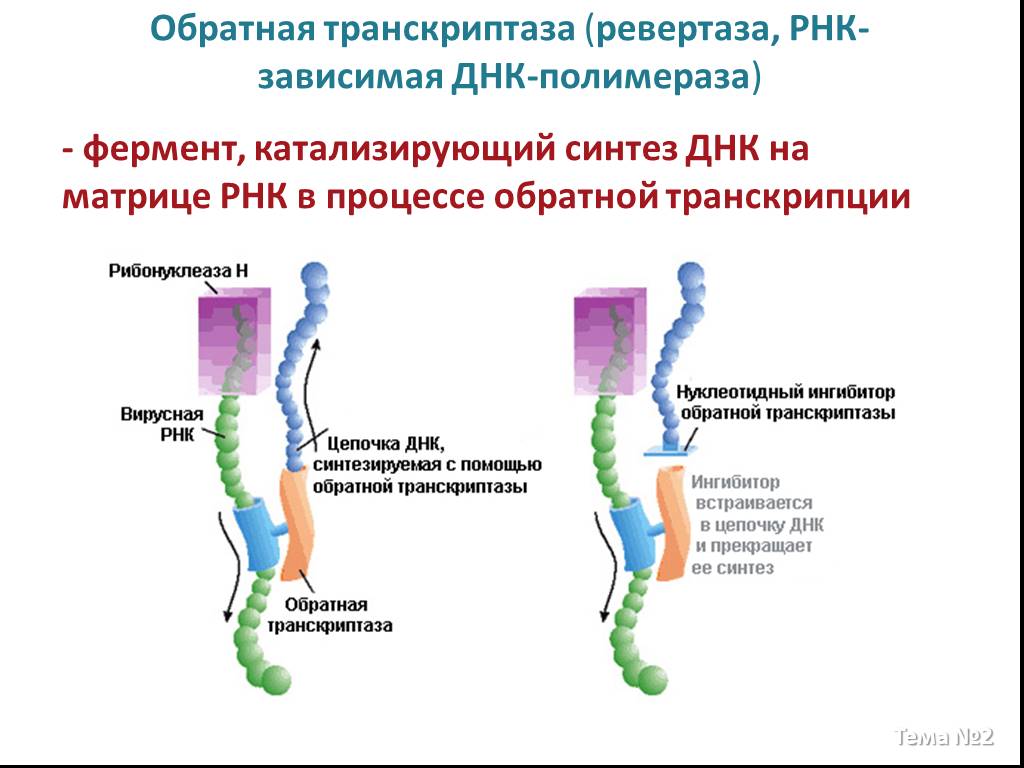 Слайд 17
Слайд 17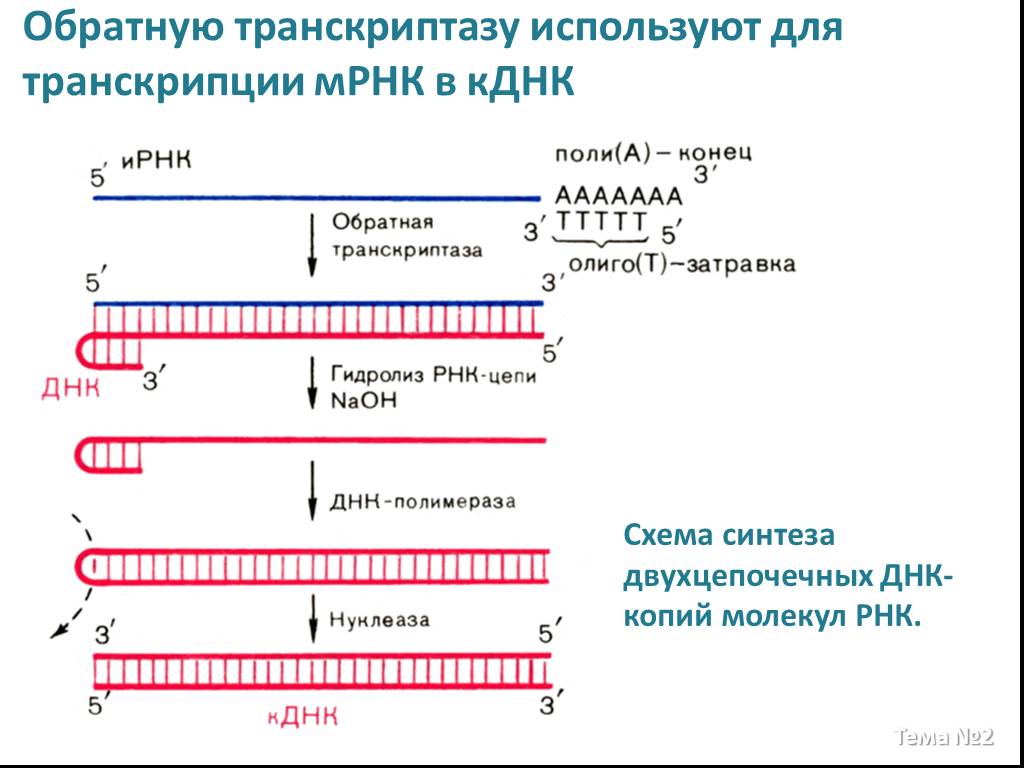 Слайд 18
Слайд 18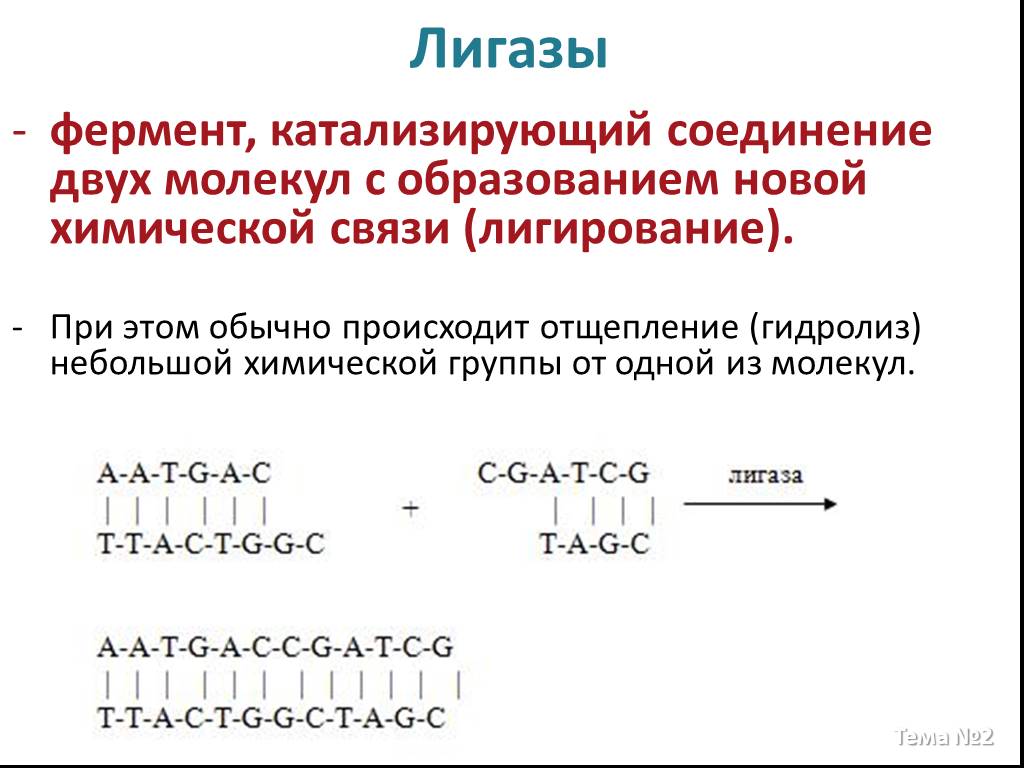 Слайд 19
Слайд 19 Слайд 20
Слайд 20 Слайд 21
Слайд 21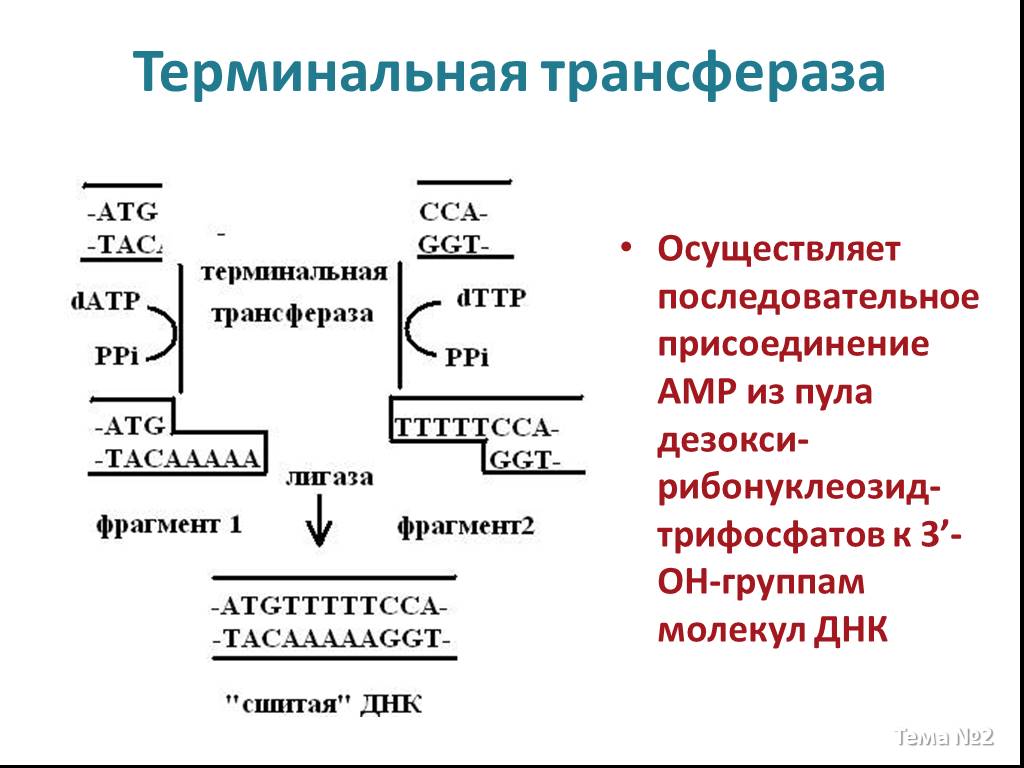 Слайд 22
Слайд 22 Слайд 23
Слайд 23 Слайд 24
Слайд 24Презентацию на тему "Основы генетической инженерии" можно скачать абсолютно бесплатно на нашем сайте. Предмет проекта: Разные. Красочные слайды и иллюстрации помогут вам заинтересовать своих одноклассников или аудиторию. Для просмотра содержимого воспользуйтесь плеером, или если вы хотите скачать доклад - нажмите на соответствующий текст под плеером. Презентация содержит 24 слайд(ов).
Слайды презентации
Список похожих презентаций

Принципы генной инженерии микроорганизмов
Генетическая инженерия (генная инженерия) — совокупность приёмов, методов и технологий получения рекомбинантных РНК и ДНК, выделения генов из организма ...
Основы туризма
«туризм — это активный отдых, влияющий на укрепление здоровья, физическое развитие человека, связанный с передвижением за пределами постоянного места ...
Основы управления персоналом банка
1. Стили руководства, их характеристика 2. Управление кадровыми резервами 3. Подсистема оплаты труда персонала коммерческого банка 4. Банковский персонал ...
Основы расчета железобетонных конструкций
бетон. Одним из основных материалов для производства ж/б конструктивных элементов является. Наиболее часто в конструкциях используют тяжелые бетоны. ...
Основы Тайм Менеджмента
? Что такое тайм менеджмент? Управление временем, т.е. технология организации времени и повышения эффективности его использования. Структурирование ...
Основы предпринимательского дела
Структура курса. Сущность предпринимательства История предпринимательства в России Предпринимательство и бизнес Виды предпринимательской деятельности ...
Основы психолого-педагогического сопровождения детей с ОВЗ в условиях инклюзивного профессионального образования
Цель: формирование у будущих бакалавров профессионального образования системы научных представлений об инклюзивном образовании лиц с ОВЗ, осуществление ...
Основы Десмургии
Общая характеристика повязок: Раздел хирургии, изучающий типы и виды повязок, правила, способы и цели их наложения, называется десмургией. Процесс ...
Основы защиты информации
План лекции. Защита информации как закономерность развития компьютерных систем Объекты и элементы защиты в компьютерных системах обработки данных ...
Основы государственной политики по организации борьбы с терроризмом
Терроризм : определение. В праве России терроризм определяется как идеология насилия и практика воздействия на общественное сознание, на принятие ...
Основы гуманистического воспитания
Традиционное обучение основано на положении учителя в качестве субъекта, а ученика - объекта педагогического процесса. В концепции сотрудничества ...
Основы видеосъемки
Крупность. Дальний. Это панорамы, пейзажи. Главный персонаж занимает около 1/3 кадра и меньше. Дальним планом можно показать общее место событий, ...
Основы архивного хранения
22 октября 2004 года № 125-ФЗ ФЕДЕРАЛЬНЫЙ ЗАКОН ОБ АРХИВНОМ ДЕЛЕ В РОССИЙСКОЙ ФЕДЕРАЦИИ Принят Государственной Думой 1 октября 2004 года Одобрен Советом ...
Основы автоматизации производственных процессов
Основы теории автоматического управления. Теория автоматического управления - наука, которая изучает процессы управления, методы их исследования и ...
Математический пакет mathcad. Основы работы с системой mathcad
Математические пакеты. Начиная с 90-х годов прошлого века, широкую известность приобрели так называемые системы компьютерной математики или математические ...
Основы землепользования и землеустройства
МОДУЛЬ 3. ОРГАНИЗАЦИОННО-ЭКОНОМИЧЕСКИЕ ОСНОВЫ ЗЕМЛЕУСТРОЙСТВА. Тема 3.3. Землеустроительная документация Вопросы: Виды землеустроительной документации ...
Основы права
Т. 1.1. Право в системе социального регулирования. Право – система, установленных государством общеобязательных правил поведения, норм, исполнение ...
Основы землепользования и землеустройства
МОДУЛЬ 3. ОРГАНИЗАЦИОННО-ЭКОНОМИЧЕСКИЕ ОСНОВЫ ЗЕМЛЕУСТРОЙСТВА. Тема 3.2. Содержание землеустроительной деятельности Вопросы: Состав землеустроительных ...
Основы прогнозирования и планирования
План лекции:. Предмет и методология курса Планирование и его этапы Прогнозирование и его классификация Принципы ПиП Факторы развития ПиП. Предмет ...
Основы землепользования и землеустройства
МОДУЛЬ 2. ГОСУДАРСТВЕННОЕ РЕГУЛИРОВАНИЕ ЗЕМЛЕПОЛЬЗОВАНИЯ. Тема 2.1. Система управления земельными ресурсами Вопросы: Принципы государственного регулирования ...Советы как сделать хороший доклад презентации или проекта
- Постарайтесь вовлечь аудиторию в рассказ, настройте взаимодействие с аудиторией с помощью наводящих вопросов, игровой части, не бойтесь пошутить и искренне улыбнуться (где это уместно).
- Старайтесь объяснять слайд своими словами, добавлять дополнительные интересные факты, не нужно просто читать информацию со слайдов, ее аудитория может прочитать и сама.
- Не нужно перегружать слайды Вашего проекта текстовыми блоками, больше иллюстраций и минимум текста позволят лучше донести информацию и привлечь внимание. На слайде должна быть только ключевая информация, остальное лучше рассказать слушателям устно.
- Текст должен быть хорошо читаемым, иначе аудитория не сможет увидеть подаваемую информацию, будет сильно отвлекаться от рассказа, пытаясь хоть что-то разобрать, или вовсе утратит весь интерес. Для этого нужно правильно подобрать шрифт, учитывая, где и как будет происходить трансляция презентации, а также правильно подобрать сочетание фона и текста.
- Важно провести репетицию Вашего доклада, продумать, как Вы поздороваетесь с аудиторией, что скажете первым, как закончите презентацию. Все приходит с опытом.
- Правильно подберите наряд, т.к. одежда докладчика также играет большую роль в восприятии его выступления.
- Старайтесь говорить уверенно, плавно и связно.
- Старайтесь получить удовольствие от выступления, тогда Вы сможете быть более непринужденным и будете меньше волноваться.
Информация о презентации
Дата добавления:17 марта 2019
Категория:Разные
Содержит:24 слайд(ов)
Поделись с друзьями:
Скачать презентацию









