Слайд 1Занятие № 4 ТЕМА:
«ВЗАИМОДЕЙСТВИЕ ГЕНОВ. ИЗМЕНЧИВОСТЬ»
Слайд 2ХРОМОСОМНАЯ ТЕОРИЯ НАСЛЕДСТВЕННОСТИ Т. МОРГАНА
Томас Гент Морган родился в 1866г. В США. В 20 лет он окончил университет, а в 25 лет стал профессором. Первоначально он был ярым противником учения Менделя. Но, занимаясь вместе со своими учениками изучением дрозофилы, он не только подтвердил открытия Менделя, но и блестяще дополнил их, создав хромосомную теорию.
 была сформулирована хромосомная теория наследственности. Гены располагаются в хромосомах, разные хромосомы содержат неодинаковое количество генов, но набор генов является строго специфичным для каждой хромосомы. Гены расположены вдоль хромосомы линейно, один")
Слайд 3Т. Морганом и его группой (1910—1916 гг.) была сформулирована хромосомная теория наследственности.
Гены располагаются в хромосомах, разные хромосомы содержат неодинаковое количество генов, но набор генов является строго специфичным для каждой хромосомы. Гены расположены вдоль хромосомы линейно, один за другим, но каждый ген находится в своем, четко определенном месте (локусе). Гены, расположенные на одной хромосоме могут передаваться потомкам совместно (сцепленно), и образуют группу сцепления. У человека 24 группы сцеплений: 22 пары аутосом, X и У-хромосомы. Сцепление не абсолютно, так как в профазе мейоза может происходить кроссинговер и гены, находящиеся в одной хромосоме, разобщаются. Сила сцепления зависит от расстояния между генами в хромосоме: чем больше расстояние, тем меньше сила сцепления, и наоборот. Расстояние между генами измеряется в % кроссинговера. 1% кроссинговера соответствует одной марганиде.

Слайд 4Знаменитый ученик Т. Моргана — Альфред Стертевант в своей дипломной работе впервые предположил, что, чем ближе друг к другу расположены гены на хромосоме, тем с большей вероятностью они будут передаваться потомкам совместно, т.е. сцепленно. Чем дальше один от другого находятся гены одной группы сцепления, тем чаще они наследуются независимо, как это следует из 3-го закона Менделя. Причиной такого независимого распределения в потомстве генов, расположенных на одной хромосоме, является кроссинговер, т.е. обмен участками хроматид между гомологичными хромосомами в профазе первого деления мейоза. В результате, кроссинговер более вероятен между генами, расположенными на хромосоме далеко друг от друга. Значительно реже он может произойти между близко находящимися генами, тогда они будут чаще наследоваться совместно.
Слайд 5
Слайд 6
Слайд 7Карты хромосом человека
Открытия Т. Моргана создали основу для определения мест расположения генов и оценки расстояния между ними. Расстояние между генами измеряется частотой кроссинговера, т.е. отношением количества особей, которые унаследовали только один ген, к числу тех, у кого были представлены совместно оба гена, выраженным в процентах. Единицей такого расстояния является 1% кроссинговера, который в честь Т.Моргана получил название 1санти-морганида (сМ).
Слайд 8Расстояние между генами в 1 сантиморганиду (1% кроссинговера) указывает, что они обычно передаются потомкам совместно (сцепленно). Если эта цифра равна 50 (50% кроссинговера), то это означает, что гены наследуются независимо друг от друга. Возможность оценки расстояния между генами стала основой для построения генетических карт хромосом.
Слайд 9ГАМЕТЫ КРОССОВЕРНЫЕ НЕКРОССОВЕРНЫЕ
Если в гамету попадает хроматида, принимавшая участие в кроссинговере (17% в опыте Моргана)
Если в гамету попадает хроматида, не принимавшая участие в кроссинговере (83% в опыте Моргана)
Слайд 10Зная расстояние между генами, можно картировать хромосомы.
Схема генетической (А) и цитологической (Б) карт хромосом
Слайд 11карты хромосом
Генетическая карта — это отрезок прямой, на котором указывается порядок расположения генов относительно друг друга и расстояние между ними в сантиморганидах. Для построения генетической карты первоначально устанавливают группы сцепления генов по анализу распределения признаков в семьях. Затем группы сцепления генов соотносят с определенными участками на хромосомах.
Цитологическая карта — это отражение ее морфологической структуры. Это или фотография, или точный рисунок хромосомы, на котором отмечается последовательность расположения генов. Ее строят на основе сопоставления результатов анализирующего скрещивания и хромосомных перестроек.
 человека составляет 3300 сМ. К настоящему времени определено расположение около 6000 локусов генома человека, которые могут наследоваться в соответствии с законами Менделя. В 1990 г. была разработана Междунар")
Слайд 12В соответствии с современными представлениями общая длина генома (совокупности всех генов) человека составляет 3300 сМ. К настоящему времени определено расположение около 6000 локусов генома человека, которые могут наследоваться в соответствии с законами Менделя. В 1990 г. была разработана Международная программа «Геном человека». Исследование генома человека открывает путь молекулярной медицине, которая обеспечит диагностику, лечение и профилактику наследственных и ненаследственных болезней с помощью генов.
Слайд 13Генетика пола
Пол — это совокупность морфологических, физиологических, биохимических, поведенческих и других признаков организма, обусловливающих репродукцию (воспроизведение себе подобных).
Слайд 14Признаки пола
Первичные половые признаки
принимают непосредственное участие в процессах воспроизведения (гаметогенез, осеменение, оплодотворение). Это наружные и внутренние половые органы. Сформированы уже к моменту рождения.
Вторичные половые признаки
не принимают непосредственного участия в репродукции, но способствуют привлечению особей разного пола. Они зависят от первичных половых признаков и развиваются под воздействием половых гормонов (у человека в 11-15 лет). Это особенности развития костно-мышечной системы, волосяного покрова, тембр голоса, поведение и др.
Слайд 15Соматические признаки особей, обусловленные полом, подразделяются на три группы: ограниченные полом, контролируемые полом и сцепленные с полом.
Слайд 16Дифференцировка пола (физикальные детерминанты пола).
Генетический пол определяется набором половых хромосом. Эмбрионы вплоть до 6 недель беременности являются потенциально бесполыми. На 6 неделе, когда закладывается индифферентная половая железа, эмбрион бисексуален. Дифференцировка эмбриональной гонады происходит у мужского организма на 7-8 неделе, а у женского — на 8-9 неделе.
Слайд 17Формирование семенников и яичников обуславливает гонадный пол, который является истинным полом. В гонадах происходит гаметогенез, что определяет гаметный пол. Кроме этого, в гонадах происходит синтез гормонов (в семенниках — тестостерона, а в яичниках — эстрогенов и прогестерона), т. е. формируется гормональный пол. В свою очередь гормоны определяют морфологический (соматический) пол: строение половой системы и развитие вторичных половых признаков. На основании морфофизиологического пола производится соответствующая запись в документах (паспорте) — гражданский пол (промежуточная детерминанта).
 полное доминирование; 2) неполное доминирование; 3)кодоминирование; 4)сверхдоминирование. Во многих случаях на проявление признака могут влиять две (или более) пары неаллельных генов. Это приводит к отклонению")
Слайд 18ВЗАИМОДЕЙСТВИЕ ГЕНОВ
ВЗАИМОДЕЙСТВИЕ АЛЛЕЛЬНЫХ ГЕНОВ
ВЗАИМОДЕЙСТВИЕ НЕАЛЛЕЛЬНЫХ ГЕНОВ
1) полное доминирование; 2) неполное доминирование; 3)кодоминирование; 4)сверхдоминирование
Во многих случаях на проявление признака могут влиять две (или более) пары неаллельных генов. Это приводит к отклонению от законов Менделя. Основные формы взаимодействия : 1)комплементарность 2) эпистаз; 3) полимерия;
Слайд 19Полное доминирование — когда один ген полностью подавляет действие другого.
Карие - А , голубые - а Р: АА х аа G: a a F1: Аа— 100 % карие глаза ВЗАИМОДЕЙСТВИЕ АЛЛЕЛЬНЫХ ГЕНОВ
Слайд 20Неполное доминирование — когда один ген не полностью подавляет другой и появляется промежуточный признак
АА — курчав волосы Р: АА х аа Аа- волнистые G: А a аа —прямые F1: Аа — 100 % волнистые
Слайд 21Кодоминирование — когда в гетерозиготном состоянии каждый аллель детерминирует свой признак.
Рассмотрим данное взаимодействие на примере АВ (IV) группы крови человека, где ген 1А определяет антиген А, ген 1в — антиген В.
Слайд 22Сверхдоминирование — когда доминантный ген в гетерозиготном состоянии проявляется сильнее, чем в гомозиготном.
Пример: продолжительность жизни у мушки дрозофилы. АА — нормальная продолжительность жизни; Аа — увеличенная продолжительность жизни; аа — летальный исход.
Слайд 23Комплементарность — взаимодействие неаллельных генов, при котором один доминантный ген дополняет действие другого доминантного гена и появляется новый признак, отсутствовавший у родителей. ПРИМЕР: Синтез интерферона у человека. Для защиты от вирусов в иммунокомпетентных клетках человека вырабатывается специфический белок интерферон. Его образование в организме связано с комплементарным взаимодействием двух неаллельных генов, локализованных в разных хромосомах.
Слайд 24Э п и с т а з (от греч. epistasis — остановка, препятствие) — взаимодействие неаллельных генов, при котором ген одной аллельной пары подавляет действие гена другой пары.
Подавляющий ген называется эпистатическим (супрессором), а подавляемый — гипостатическим. В случаях, когда ген-подавитель доминантный, имеет место эпистаз.
Слайд 25КРИПТОМЕРИЯ
В случаях, когда ген-подавитель рецессивный, имеет место криптомерия греч. kryptos — тайный, скрытый). У человека примером криптомерии может служить «бомбейский феномен», в этом случае редкий рецессивный аллель «х» в гомозиготном состоянии «хх » подавляет активность гена 1В (который детерминирует В(III) группу крови системы АВО), поэтому женщина, получившая от матери аллель 1В, фенотипически имела первую группу крови — 0(1).
 — когда несколько доминантных генов определяют один и тот же признак примерно в одинаковых количествах. С помощью полимерных генов наследуются количественные признаки. Чем больше число доминантных полимерных генов, сильнее выражен признак. У человека п")
Слайд 26Полимерия (от греч. polymeria — многосложность) — когда несколько доминантных генов определяют один и тот же признак примерно в одинаковых количествах.
С помощью полимерных генов наследуются количественные признаки. Чем больше число доминантных полимерных генов, сильнее выражен признак. У человека полимерными генами определяется пигментация кожи. У коренных жителей Африки (негроидной расы) преобладают доминантные аллели- А, у представителей европеоидной — рецессивные- а. Мулаты имеют промежуточную пигментацию. Чем больше доминантных генов в генотипе, тем темнее кожа, и наоборот, чем меньше, тем она светлее. Дети от брака негра и светлой женщины будут мулатами (А1а1 А2а2). В потомстве двух мулатов вероятность рождения негра (А1А1 А2А2) или белого (а1а1 а2а2) равна 1/16. Остальные генотипы попадают в промежуточные фенотипические классы.
. ГЕНОТИПИЧЕСКАЯ (наследственная). КОМБИНАТИВНАЯ МУТАЦИОННАЯ")
Слайд 27ИЗМЕНЧИВОСТЬ — свойство организмов приобретать в процессе онтогенеза отличия признаков от родительских форм.
Изменчивость
ФЕНОТИПИЧЕСКАЯ (модификационная, ненаследственная)
ГЕНОТИПИЧЕСКАЯ (наследственная)
КОМБИНАТИВНАЯ МУТАЦИОННАЯ
Слайд 28Фенотипическая (модификационная) изменчивость — изменения фенотипа под действием факторов внешней среды без изменения структуры генотипа.
Фенотипическая изменчивость была открыта Бонне. Корневую систему 120 видов растений он разделял на 2 части, одну часть высаживал в Парижском ботаническом саду, другую — в горах. В разных условиях вырастали растения с разными фенотипами (высокие и низкие). Бонне собрал семена таких растений и высадил в одинаковых условиях — выросли одинаковые растения.
Слайд 29Свойства модификаций:
ненаследуемость; приспособительный (адаптивный) характер изменений; определенность (направленность и предсказуемость изменений, возникающих в организме под действием фактора среды); степень изменения признака под действием факторов среды зависит от времени и силы его действия; носит групповой характер; не является материалом для естественного отбора.
Слайд 30Норма реакции — пределы модификационной изменчивости, которые формируются на основе генотипа в разных условиях внешней среды.
Норма реакции
ШИРОКАЯ (масса тела)
УЗКАЯ (цвет волос)
Фенокопии — явление, когда ненаследственная изменчивость копирует наследственную изменчивость. Например: у европеоида(с белой кожей) при продолжительном воздействии УФ кожа становится пигментированной и копирует цвет кожи монголоида, у которого другой генотип
Слайд 31Комбинативная изменчивость — появление новых признаков у потомства, вследствие возникновения новых комбинаций генов родителей при образовании и слиянии их гамет.
Механизмы комбинативной изменчивости: Кроссинговер (профаза мейоза I); Независимое расхождение хромосом в анафазу мейоза I; Независимое расхождение хроматид в анафазу мейоза И; Случайное сочетание гамет при оплодотворении.
Слайд 32Мутационная изменчивость (мутация) — скачкообразное и устойчивое изменение генетического материала, передающееся по наследству.
Свойства мутаций: « возникают внезапно, скачкообразно; наследуются; ненаправленны (неопределенны), т. е. под действием одного фактора может мутировать любой ген; в основном, являются вредными для организма; индивидуальны; являются материалом для действия естественного отбора; постоянны.
Слайд 33Генокопии — одинаковое фенотипическое проявление мутаций разных генов. Пример генокопий — различные виды гемофилии, связанные с недостаточностью VIII и IX факторов свертывающей системы (гемофилия а и в). Мутагенез - процесс образования мутаций Мутагены - факторы, вызывающие мутации
Слайд 34Классификация мутагенных факторов:
Физические (рентгеновское и ионизирующее излучения, температура, УФЛ др.); Химические (формалин, нитриты, нитраты, аналоги азотистых оснований, иприт, лекарства, алкоголь, никотин и др.); Биологические (вирусы, бактерии, токсины паразитов). Эти факторы вызывают разнообразные изменения генетического материала: разрывы в молекулах ДНК; Т-Т димеры в молекулах ДНК; разрушают нити веретена деления; нарушают процесс кроссинговера; приводят к образованию в организме свободных радикалов.
Слайд 35Классификация мутаций:
Слайд 36По исходу для организма мутации бывают:
отрицательные — летальные (несовместимы с жизнью); полулетальные (снижают жизнеспособность организма); нейтральные (не влияют на жизнедеятельность); • положительные (повышают жизнеспособность) — возникают редко, но имеют большое значение для эволюции.
Слайд 37По изменениям генетического материала:
Геномные мутации - обусловлены изменениями числа хромосом. Хромосомные мутации (аберрации) - изменения структуры хромосом. Генные мутации (трансгенации) - изменения структуры гена.
. Полиплоидия используется в селекции растений и приводит к повышению урожайности. У млекопитающих и человека это летальные мутации. Гаплоидия (1n) — одинарный набо")
Слайд 38ГЕНОМНЫЕ МУТАЦИИ
Всегда проявляются фенотипически. Полиплоидия — кратное гаплоидному набору увеличение числа хромосом (Зп, 4п, 5п, ...). Полиплоидия используется в селекции растений и приводит к повышению урожайности. У млекопитающих и человека это летальные мутации. Гаплоидия (1n) — одинарный набор хромосом, например, у трутней пчел. Жизнеспособность гаплоидов снижается, так как у них проявляются все рецессивные гены, содержащиеся в единственном числе. Для млекопитающих и человека — летальная мутация. Анеуплоидия — некратное гаплоидному уменьшение или увеличение числа хромосом (2п±1, 2п±2, и т. д.). Разновидности анеуплоидии: • трисомия — 3 гомологичных хромосомы в кариотипе (2п+1): - синдром трисомии X. Частота встречаемости 1:800-1:1000. Кариотип 47, XXX; - синдром Патау. Частота — 1:6000. Кариотип 47, XX, 13+ или 47 ХУ, 13+; - синдром Эдвардса. Частота — 1:7000. Кариотип 47, XX, 18+ или 47 ХУ, 18+; - синдром Дауна. Частота — 1:900. Кариотип 47, XX, 21+ или 47, ХУ, 21+; - синдром Кляйнфельтера. Частота — 1:400-1:500. Кариотип 47, ХХУ; 48, ХХХУ; • моносомия — в диплоидном наборе одна из пары гомологичных хромосом: - синдром Шерешевского-Тернера: Частота— 1:2000-1:3000. Кариотип 45, Х0. Моносомии по первым крупным парам хромосом для человека — летальные мутации. • нулисомия — отсутствие пары хромосом в кариотипе (летальная мутация).
 — выпадение части хромосомы Пр: делеция участка короткого плеча 5-й хромосомы у человека (5р-) — синдром кошачьего крика. дупликация — удвоение участка хромосомы. инверсия — отрыв участка хромосомы, пово")
Слайд 392. К внутрихромосомным мутациям относятся перестройки внутри одной хромосомы:
делеция (нехватка) — выпадение части хромосомы Пр: делеция участка короткого плеча 5-й хромосомы у человека (5р-) — синдром кошачьего крика. дупликация — удвоение участка хромосомы. инверсия — отрыв участка хромосомы, поворот его на 180° и прикрепление к месту отрыва. При этом наблюдается нарушение порядка расположения генов. Делеции и дупликации проявляются фенотипически всегда, так как изменяется набор генов и наблюдаются частичные моносомии при нехватках и частичные трисомии — при дупликациях.
; - нереципрокные (сегменты одной хромосомы переносятся в другую); - робертсоновские (")
Слайд 402. Межхромосомные мутации происходят между негомологичными хромосомами.
Транслокация — обмен сегментами между негомологичными хромосомами. Различают транслокации: - реципрокные (2 хромосомы обмениваются сегментами); - нереципрокные (сегменты одной хромосомы переносятся в другую); - робертсоновские (две акроцентрические хромосомы соединяются своими центромерными районами). Инверсии и транслокации фенотипически проявляются не всегда, они могут быть сбалансированными, когда не происходит ни увеличения, ни уменьшения генетического материала и сохраняется общий баланс генов в геноме.
 — изменения структуры гена. Мутации структурных генов: «сдвиг рамки считывания» — вставка или выпадение нуклеотидов (в зависимости от места вставки или выпадения изменяется меньшее или большее число кодонов); транзиция — замена оснований пуринового на пуриновое, или")
Слайд 413. Генные мутации (трансгенации) — изменения структуры гена
Мутации структурных генов: «сдвиг рамки считывания» — вставка или выпадение нуклеотидов (в зависимости от места вставки или выпадения изменяется меньшее или большее число кодонов); транзиция — замена оснований пуринового на пуриновое, или пиримидинового на пиримидиновое, например: А-Г, Ц-Т; при этом изменяется тот кодон, в котором произошла транзиция; трансверзия — замена пуринового основания на пиримидиновое или пиримидинового на пуриновое. Например: А-Ц, Г-Т; изменяется тот кодон, в котором произошла трансверзия. Изменения структурных генов приводят: к мисценс-мутациям — изменению смысла кодонов и образованию других белков; к нонсенс-мутациям — образованию «бессмысленных» кодонов (АТТ, АТЦ, АЦТ в ДНК; УАА, УАГ, УГА в и-РНК), не кодирующих АК (терминаторы, определяющие окончание считывания). Мутации функциональных генов: • белок-репрессор «не подходит» к гену-оператору («ключ не входит в замочную скважину») — структурные гены работают постоянно и белки синтезируются все время; • белок-репрессор плотно «присоединяется» к гену-оператору и не снимается индуктором («ключ не выходит из замочной скважины») — структурные гены постоянно не работают и не происходит синтез белков, закодированных в данном транскриптоне; Генные мутации в большинстве случаев проявляются фенотипически и являются причиной нарушения обмена веществ (генных болезней), частота проявления которых в популяциях человека 2-4 %.
 Слайд 1
Слайд 1 Слайд 2
Слайд 2 Слайд 3
Слайд 3 Слайд 4
Слайд 4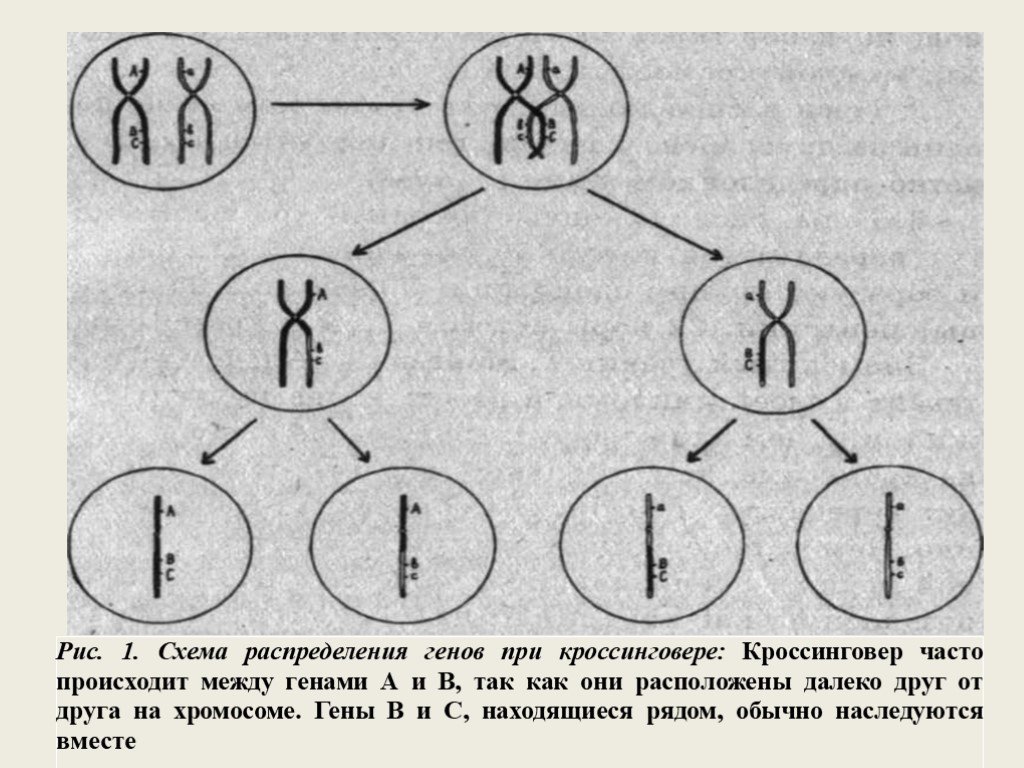 Слайд 5
Слайд 5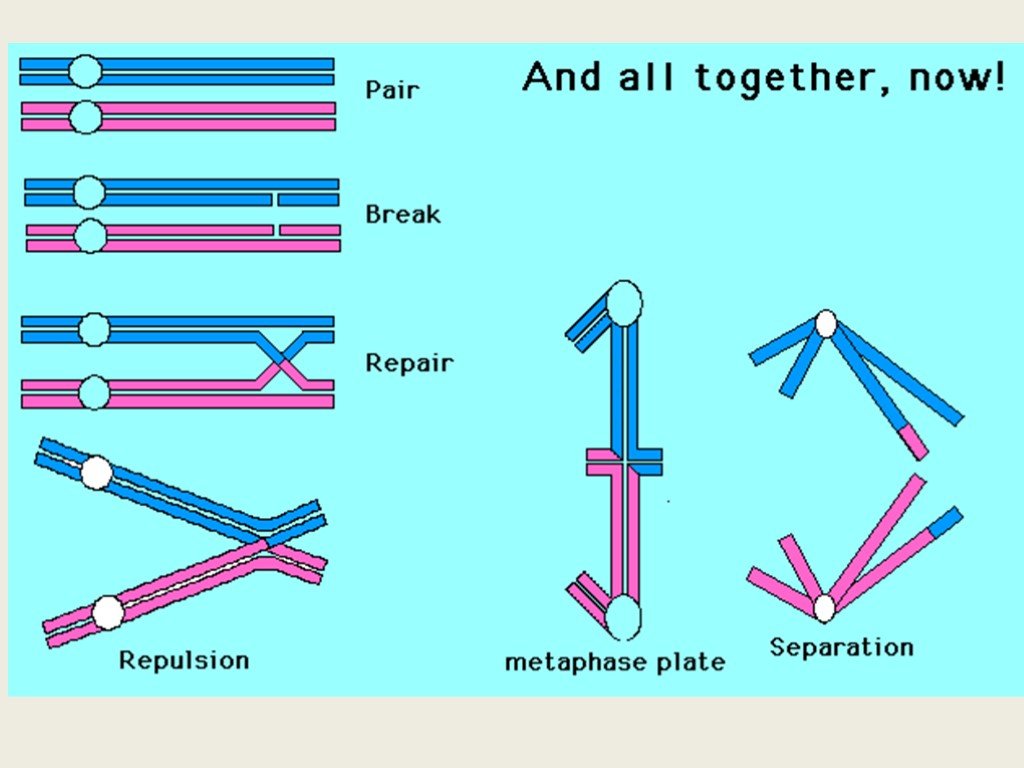 Слайд 6
Слайд 6 Слайд 7
Слайд 7 Слайд 8
Слайд 8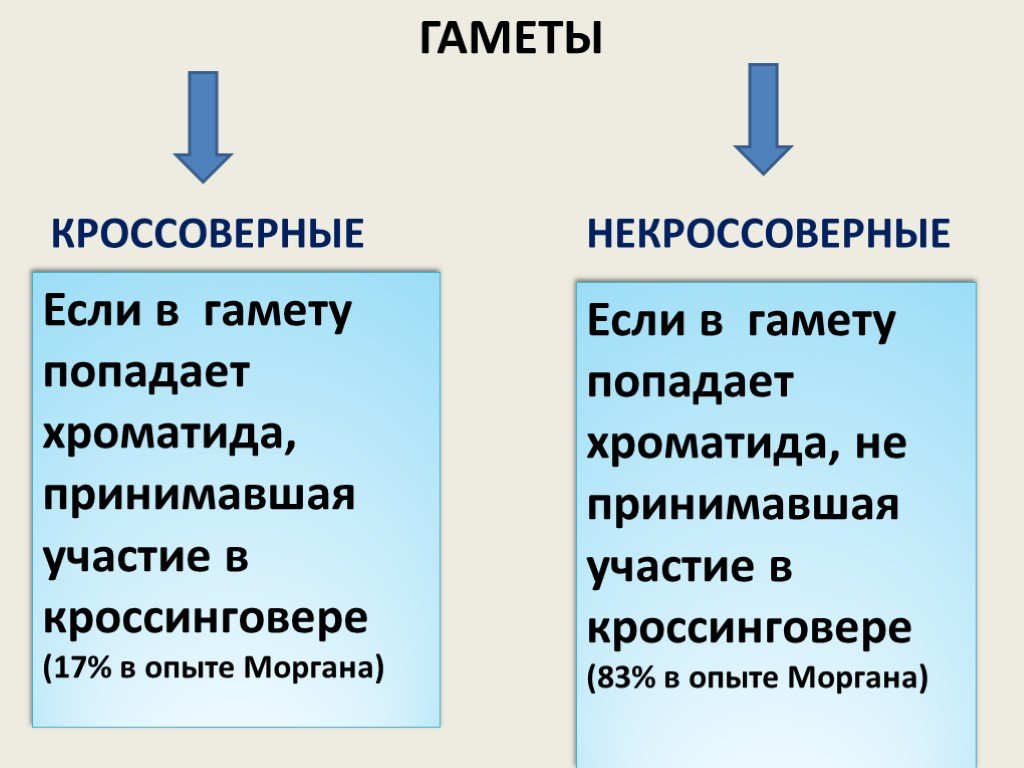 Слайд 9
Слайд 9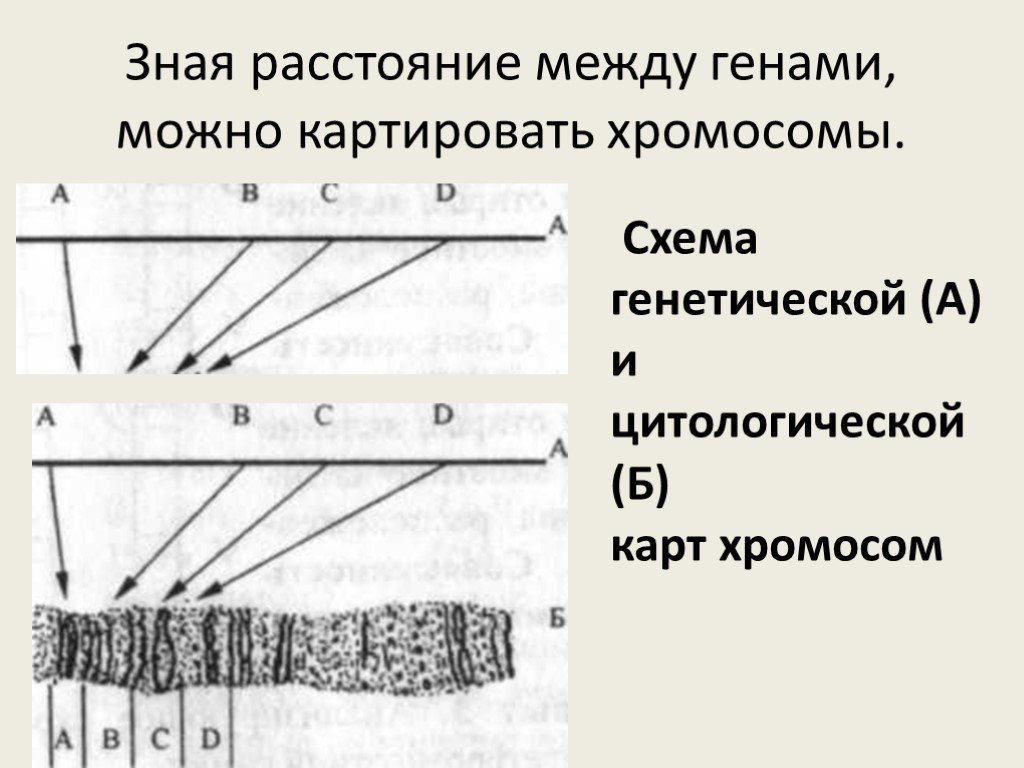 Слайд 10
Слайд 10 Слайд 11
Слайд 11 Слайд 12
Слайд 12 Слайд 13
Слайд 13 Слайд 14
Слайд 14 Слайд 15
Слайд 15 Слайд 16
Слайд 16 Слайд 17
Слайд 17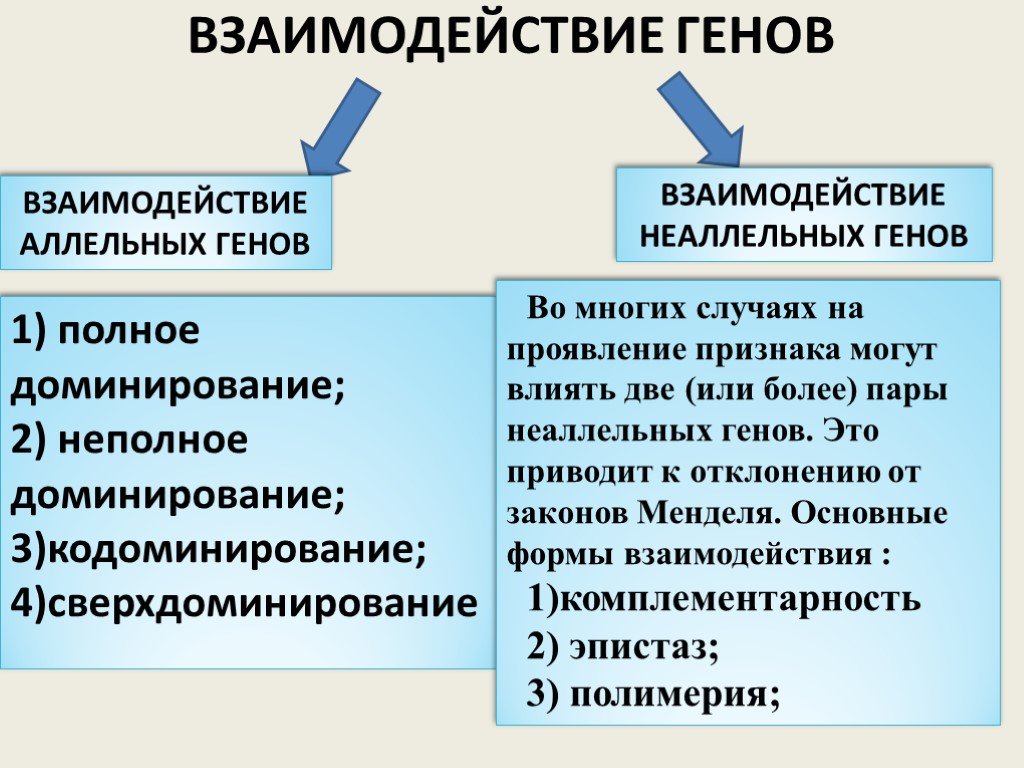 Слайд 18
Слайд 18 Слайд 19
Слайд 19 Слайд 20
Слайд 20 Слайд 21
Слайд 21 Слайд 22
Слайд 22 Слайд 23
Слайд 23 Слайд 24
Слайд 24 Слайд 25
Слайд 25 Слайд 26
Слайд 26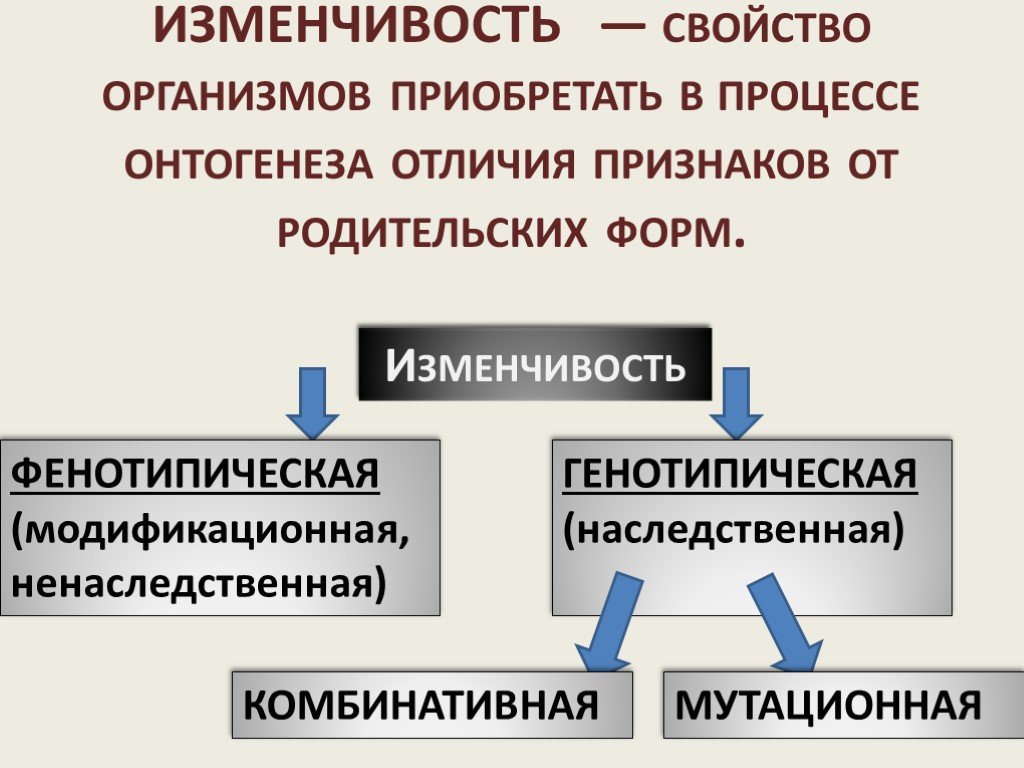 Слайд 27
Слайд 27 Слайд 28
Слайд 28 Слайд 29
Слайд 29 Слайд 30
Слайд 30 Слайд 31
Слайд 31 Слайд 32
Слайд 32 Слайд 33
Слайд 33 Слайд 34
Слайд 34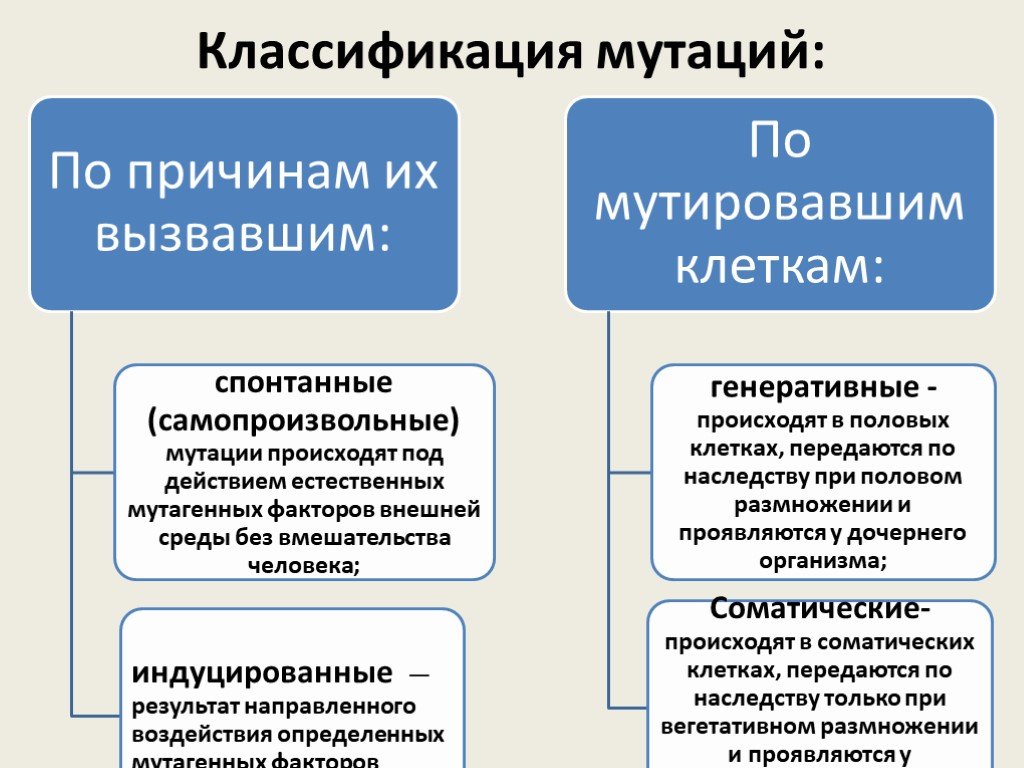 Слайд 35
Слайд 35 Слайд 36
Слайд 36 Слайд 37
Слайд 37 Слайд 38
Слайд 38 Слайд 39
Слайд 39 Слайд 40
Слайд 40 Слайд 41
Слайд 41















