Презентация "Физиология скелетных мышц" по медицине – проект, доклад
 Слайд 1
Слайд 1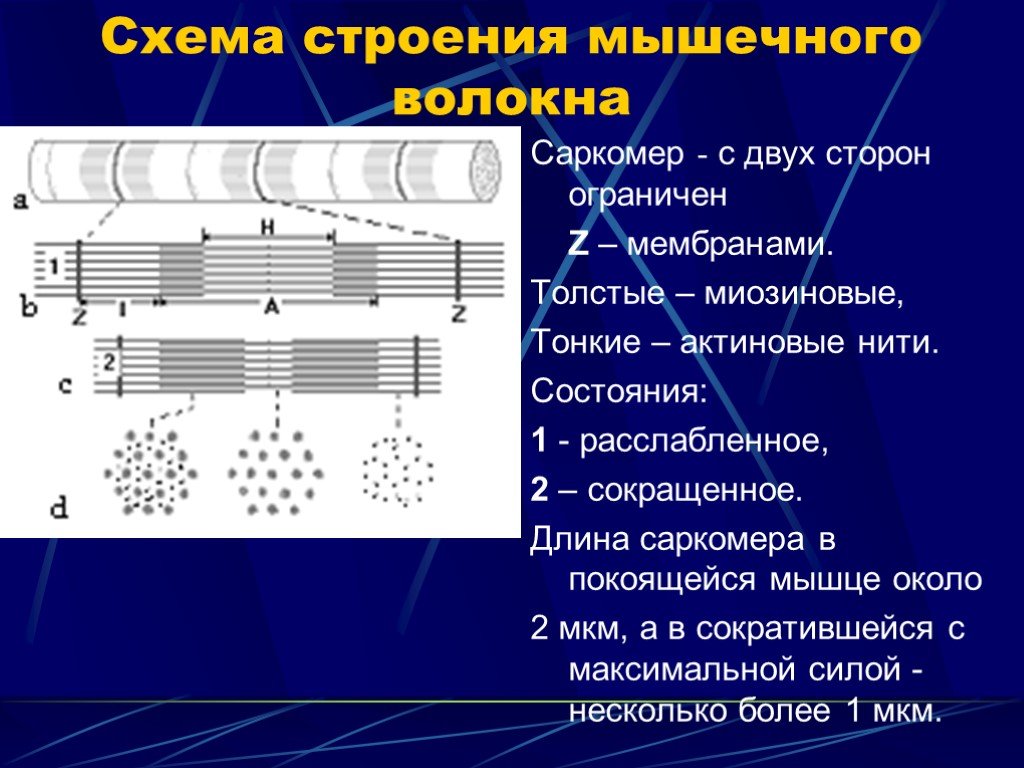 Слайд 2
Слайд 2 Слайд 3
Слайд 3 Слайд 4
Слайд 4 Слайд 5
Слайд 5 Слайд 6
Слайд 6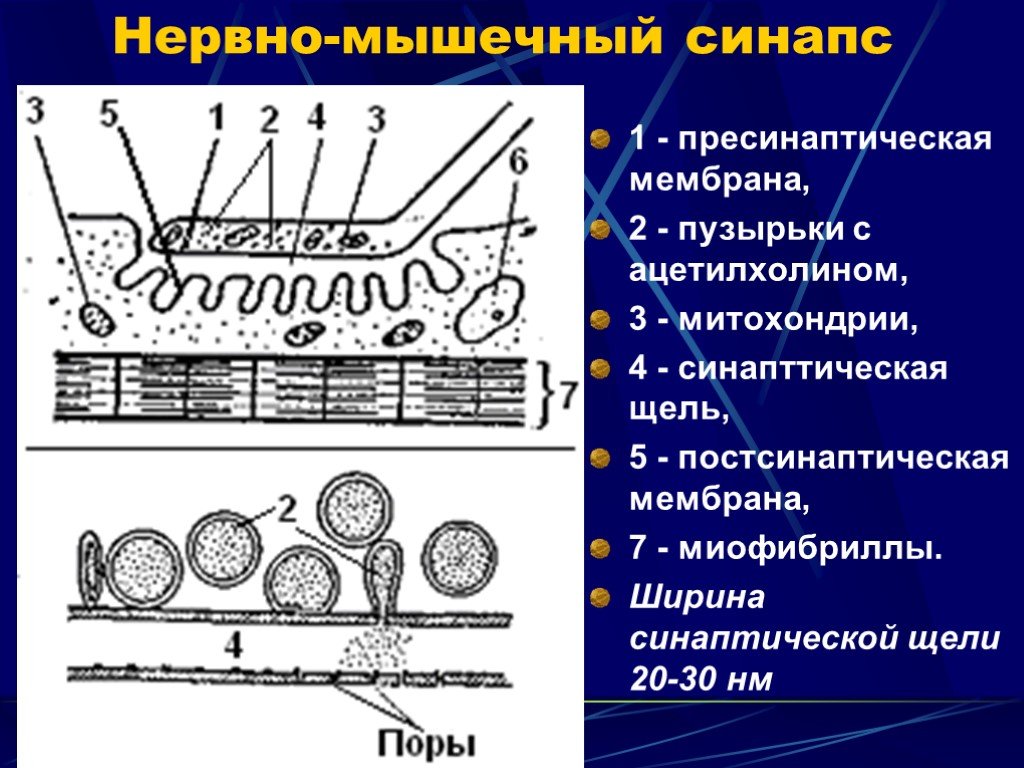 Слайд 7
Слайд 7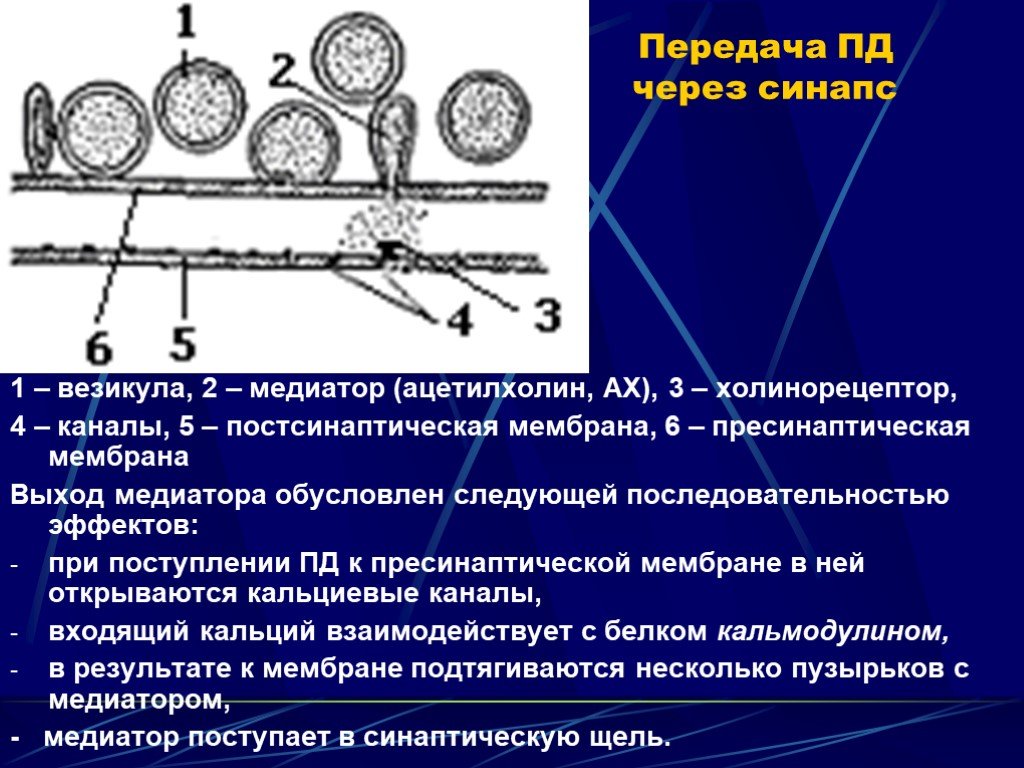 Слайд 8
Слайд 8 Слайд 9
Слайд 9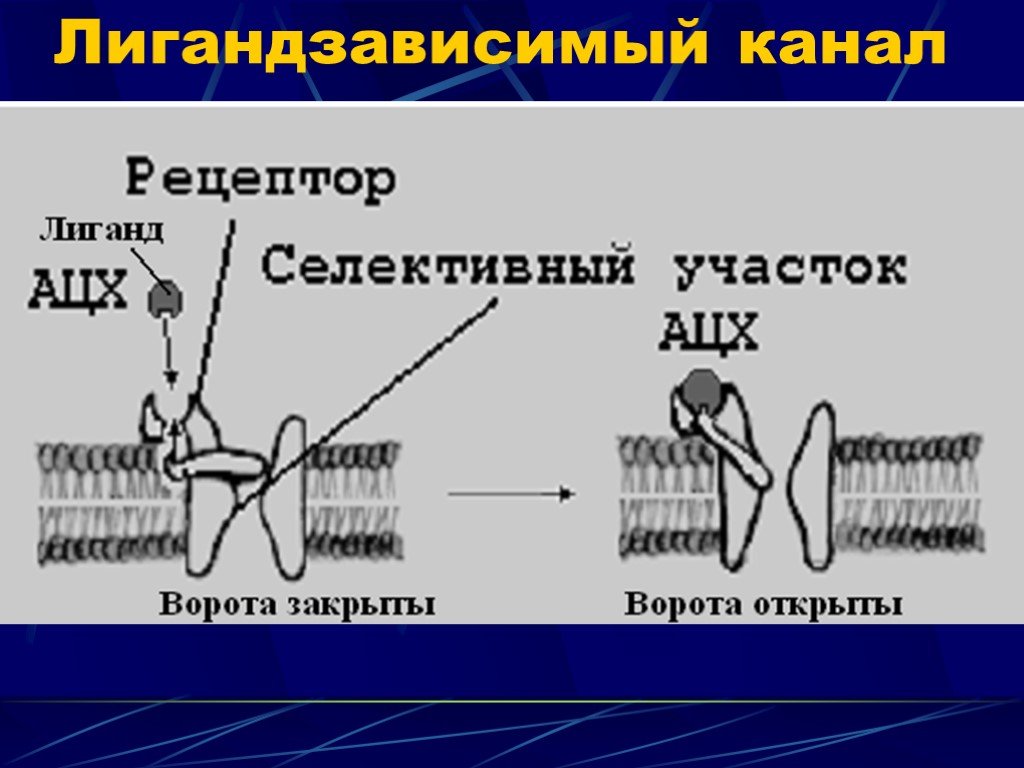 Слайд 10
Слайд 10 Слайд 11
Слайд 11 Слайд 12
Слайд 12 Слайд 13
Слайд 13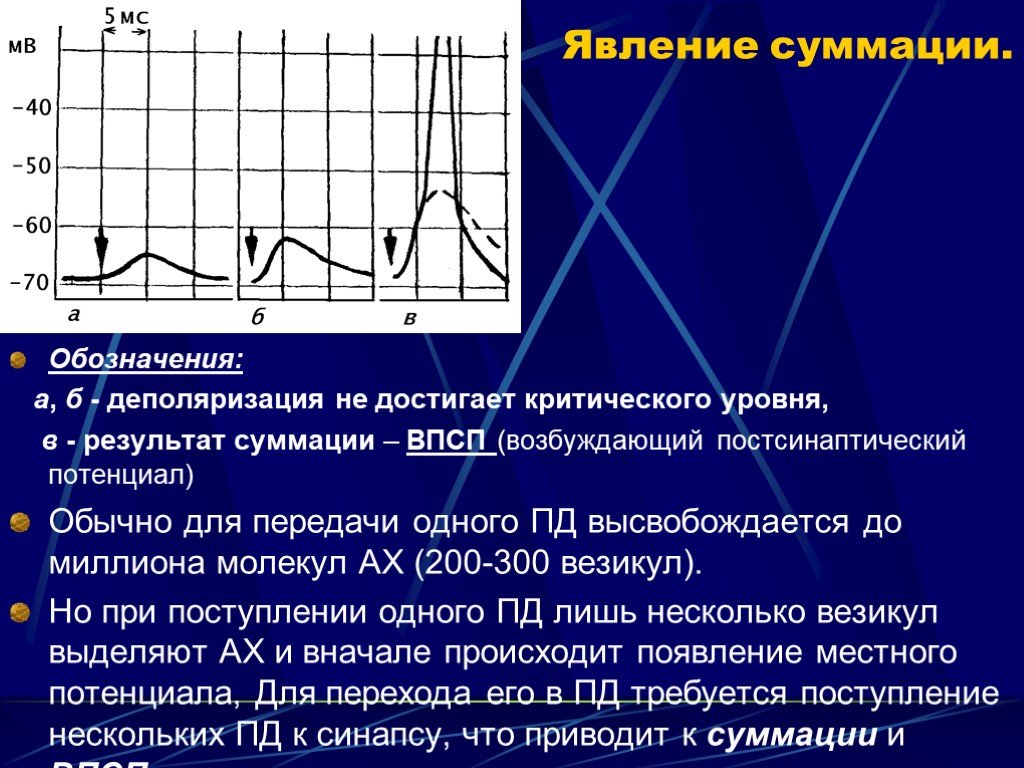 Слайд 14
Слайд 14 Слайд 15
Слайд 15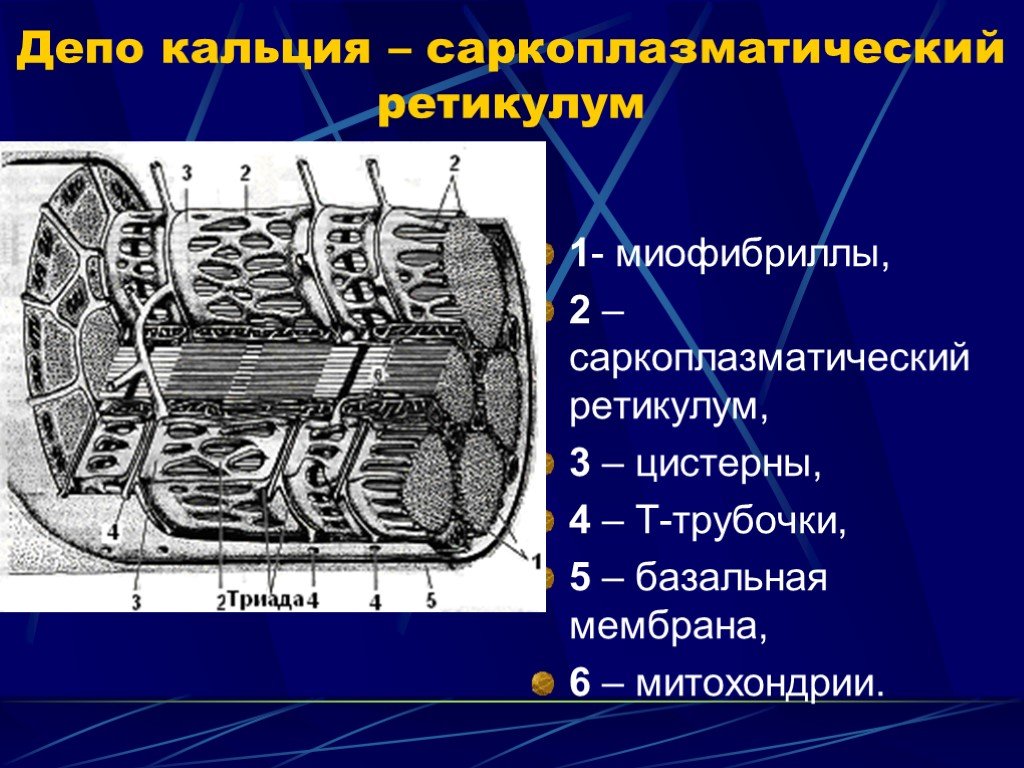 Слайд 16
Слайд 16 Слайд 17
Слайд 17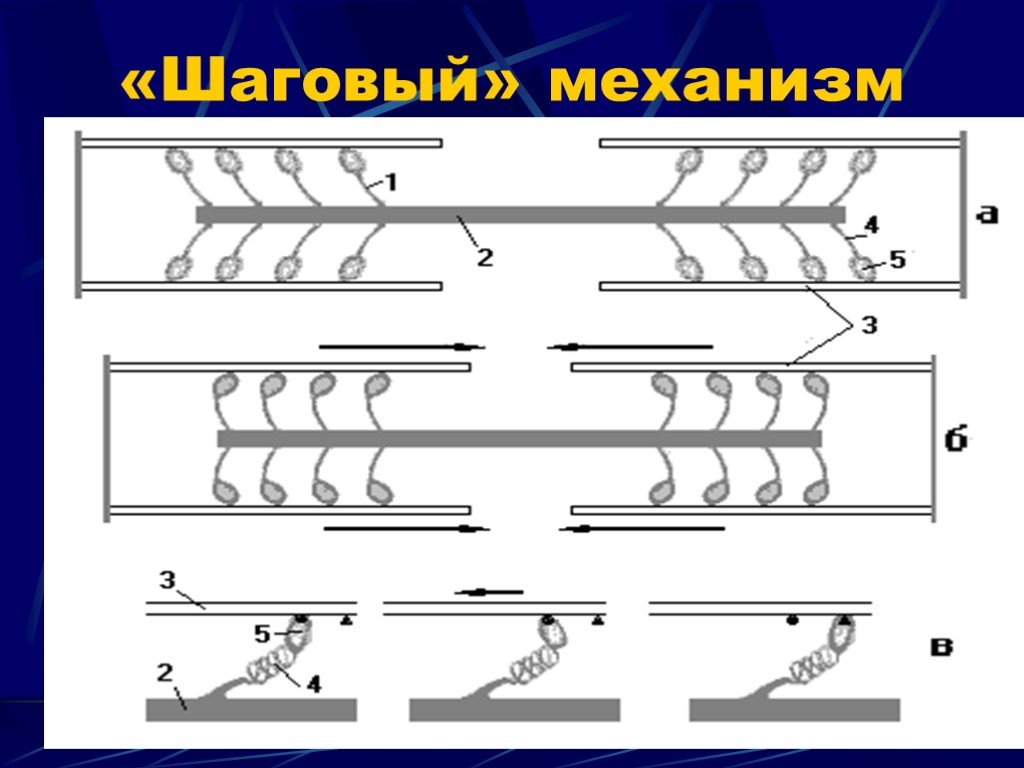 Слайд 18
Слайд 18 Слайд 19
Слайд 19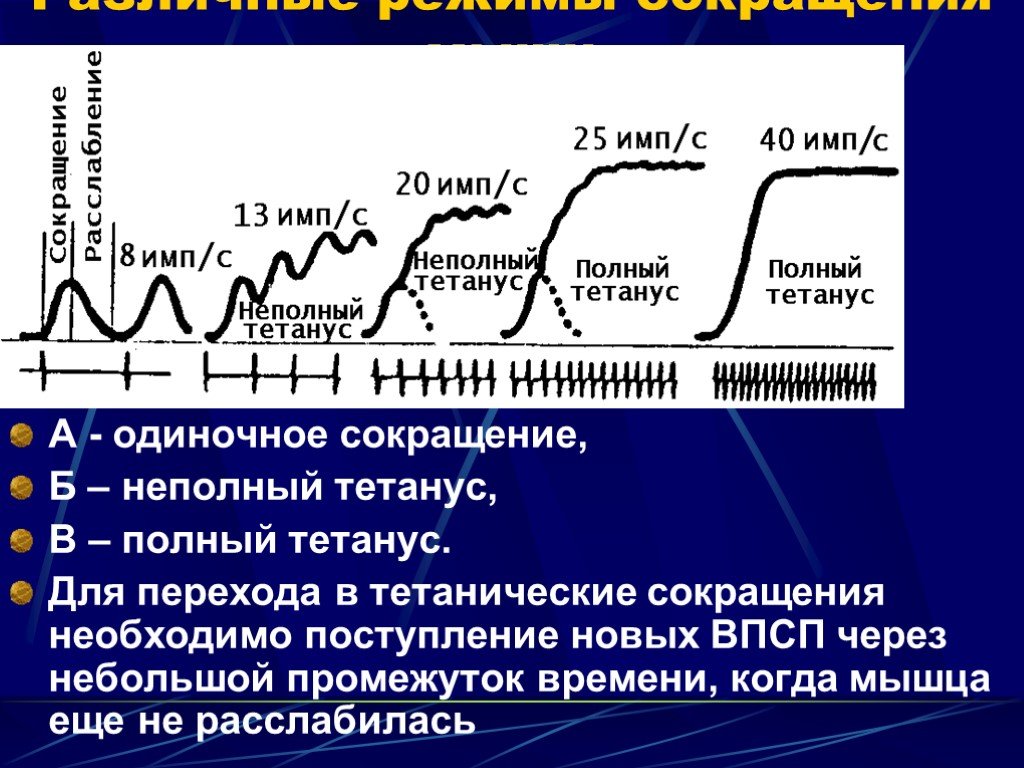 Слайд 20
Слайд 20 Слайд 21
Слайд 21 Слайд 22
Слайд 22 Слайд 23
Слайд 23 Слайд 24
Слайд 24 Слайд 25
Слайд 25 Слайд 26
Слайд 26 Слайд 27
Слайд 27 Слайд 28
Слайд 28Презентацию на тему "Физиология скелетных мышц" можно скачать абсолютно бесплатно на нашем сайте. Предмет проекта: Медицина. Красочные слайды и иллюстрации помогут вам заинтересовать своих одноклассников или аудиторию. Для просмотра содержимого воспользуйтесь плеером, или если вы хотите скачать доклад - нажмите на соответствующий текст под плеером. Презентация содержит 28 слайд(ов).
Слайды презентации
Список похожих презентаций

Физиология сокращения мышц
Иннервация мышц. Режимы мышечных сокращений. Мышечное веретено. Состояние мышечных веретен и их импульсация в разных условиях. ЭМГ при разной силе ...
Холинергическая вазодилятация сосудов скелетных мышц
Раздражение защитной области гипоталламуса. Активация симпатической нервной системы. Вазодилятация мелких сосудов скелетных мышц. Десимпатизация Атропин. ...
Физиология сна
Что такое сон? Сон - физиологическое состояние, которое характеризуется потерей активных психических связей субъекта с окружающим его миром. Сон является ...
Физиология пищеварения. Нормы питания
ЗАДАЧИ ФИЗИОЛОГИИ ПИТАНИЯ:. - оценить калорийность пищи. - сбалансировать рацион питания. - выбрать режим питания. Внешняя среда Организм Внешняя ...
Физиология почек и водно солевого обмена
ОРГАНЫ ВЫДЕЛЕНИЯ. СИСТЕМА ВЫДЕЛЕНИЯ. КОЖА - 300-1000 мл пота. 1/3 экскретируемой воды, до 10 % мочевины ЛЕГКИЕ - 400-1000 мл воды КИШЕЧНИК - до 100 ...
Физиология питания и пищеварения
ЕСТЕСТВЕННОЕ ИСКУССТВЕННОЕ ГИСТОТРОФНОЕ ДЕФИНИТИВНОЕ ИЛИ ЭНТЕРАЛЬНОЕ АМНИОТРОФНОЕ ГЕТЕРОТРОФНОЕ ГЕМОТРОФНОЕ ПАРЭНТЕРАЛЬНОЕ ЛАКТОТРОФНОЕ. ТИПЫ ПИТАНИЯ. ...
Физиология пищеварения в тонком кишечнике
ТРИ ЗВЕНА ПИЩЕВАРЕНИЯ В ТОНКОЙ КИШКЕ. Полостной гидролиз Мембранный гидролиз Всасывание. Последовательность процессов пищеварительного конвейера. ...
Физиология анализаторов
Структура анализаторной системы. Этапы деятельности анализатора. Строение глаза. Мышцы глаза. А – вид спереди, Б – вид сверху. Зрительные пути. Механизм ...
Роды Подготовка к родам Физиология родовой деятельности
ОЦЕНКА ГОТОВНОСТИ ОРГАНИЗМА БЕРЕМЕННОЙ К РОДАМ. ФИЗИОЛОГИЯ РОДОВОЙ ДЕЯТЕЛЬНОСТИ. ПРИНЦИПЫ ВЕДЕНИЯ НОРМАЛЬНЫХ РОДОВ. ПАРТОГРАММА. По определению ВОЗ, ...
Физиология иммунной системы
Иммунная система – морфофункциональная, самоорганизующаяся защитная система организма, направленная на сохранение жизнедеятельности и биологической ...
Физиология эндокринной системы
Железы внутренней секреции и их гормоны. Эндокринная система. 1. Эндокринные железы ГИПОФИЗ (аденогипофиз и нейрогипофиз) НАДПОЧЕЧНИКИ (кора и мозговое ...
Физиология гепато-билиарной системы
Схема строения гепато-билиарной системы. Печень (поддержание гомеостаза, кроветворение, выработка желчи). Желчный пузырь (накопление желчи). Общий ...
Физиология дыхания
План:. Функциональные показатели системы дыхания Газообмен Регуляция дыхания Изменение показателей дыхания при мышечных нагрузках. 1.Функциональные ...
Физиология вегетативной нервной системы
Цитата. « Мы не являемся хозяевами, а лишь свидетелями частоты сердцебиений, сокращений желудка и кишечника. Их работа совершается помимо нашей воли. ...
Физиология вегетативной нервной системы
Вегетативной нервной системе принадлежит ведущая роль в осуществлении приспо- собительных реакций организма при охлаждении, кровопотере, интенсивной ...
Физиология базальных ганглиев
Синонимы термина БГ. - подкорковые ганглии; - базальные ганглии; - стриопаллидарная система. Структуры БЯ. 1. полосатое тело (хвостатое ядро и скорлупа); ...
Физиология сна и сновидений
Что такое сон? Сон - физиологическое состояние, которое характеризуется потерей активных психических связей субъекта с окружающим его миром. Сон является ...
Физиология энергетического обмена
Общее представление об обмене веществ и энергии. Методы исследования обмена энергии. Принцип работы метаболиметра. Камера Шатерникова для человека. ...
Физиология лейкоцитов Иммунитет
Лейкоциты. Общее количество: 4,5 - 9 . 109 / л 4500-9000 в 1 мкл 2 КЛАССА: ГРАНУЛОЦИТЫ ( нейтрофилы, базофилы, эозинофилы) и АГРАНУЛОЦИТЫ ( лимфоциты ...
Топография мышц
Значение знания топографии мышц для практической медицины. 1. Топографическая анатомия составляет основу клинической анатомии 2. Изучает точное положение ...Конспекты
Строение и функции скелетных мышц
Урок 11. Строение и функции скелетных мышц. Задачи урока:. сформировать у учащихся представления о мышцах как части опорно-двигательной системы;. ...Работа скелетных мышц и их регуляция
Конспект. . урока биологии в 8 классе. . "Работа скелетных мышц и их регуляция". Учитель биологии высшей категории Емельянова В.Н. Цель:. ...Скелет поясов и свободных конечностей. Первая помощь при травмах скелета и мышц
Конкур. с. : «Лучшая методическая разработка урока с использованием ИКТ». НОМИНАЦИЯ:. Физика, математика, химия, биология, информатика. Учебный ...Советы как сделать хороший доклад презентации или проекта
- Постарайтесь вовлечь аудиторию в рассказ, настройте взаимодействие с аудиторией с помощью наводящих вопросов, игровой части, не бойтесь пошутить и искренне улыбнуться (где это уместно).
- Старайтесь объяснять слайд своими словами, добавлять дополнительные интересные факты, не нужно просто читать информацию со слайдов, ее аудитория может прочитать и сама.
- Не нужно перегружать слайды Вашего проекта текстовыми блоками, больше иллюстраций и минимум текста позволят лучше донести информацию и привлечь внимание. На слайде должна быть только ключевая информация, остальное лучше рассказать слушателям устно.
- Текст должен быть хорошо читаемым, иначе аудитория не сможет увидеть подаваемую информацию, будет сильно отвлекаться от рассказа, пытаясь хоть что-то разобрать, или вовсе утратит весь интерес. Для этого нужно правильно подобрать шрифт, учитывая, где и как будет происходить трансляция презентации, а также правильно подобрать сочетание фона и текста.
- Важно провести репетицию Вашего доклада, продумать, как Вы поздороваетесь с аудиторией, что скажете первым, как закончите презентацию. Все приходит с опытом.
- Правильно подберите наряд, т.к. одежда докладчика также играет большую роль в восприятии его выступления.
- Старайтесь говорить уверенно, плавно и связно.
- Старайтесь получить удовольствие от выступления, тогда Вы сможете быть более непринужденным и будете меньше волноваться.
Информация о презентации
Дата добавления:28 января 2019
Категория:Медицина
Содержит:28 слайд(ов)
Поделись с друзьями:
Скачать презентацию









