Презентация "Моделирование регуляции развития меристемы побега" (9 класс) по биологии – проект, доклад
 Слайд 1
Слайд 1 Слайд 2
Слайд 2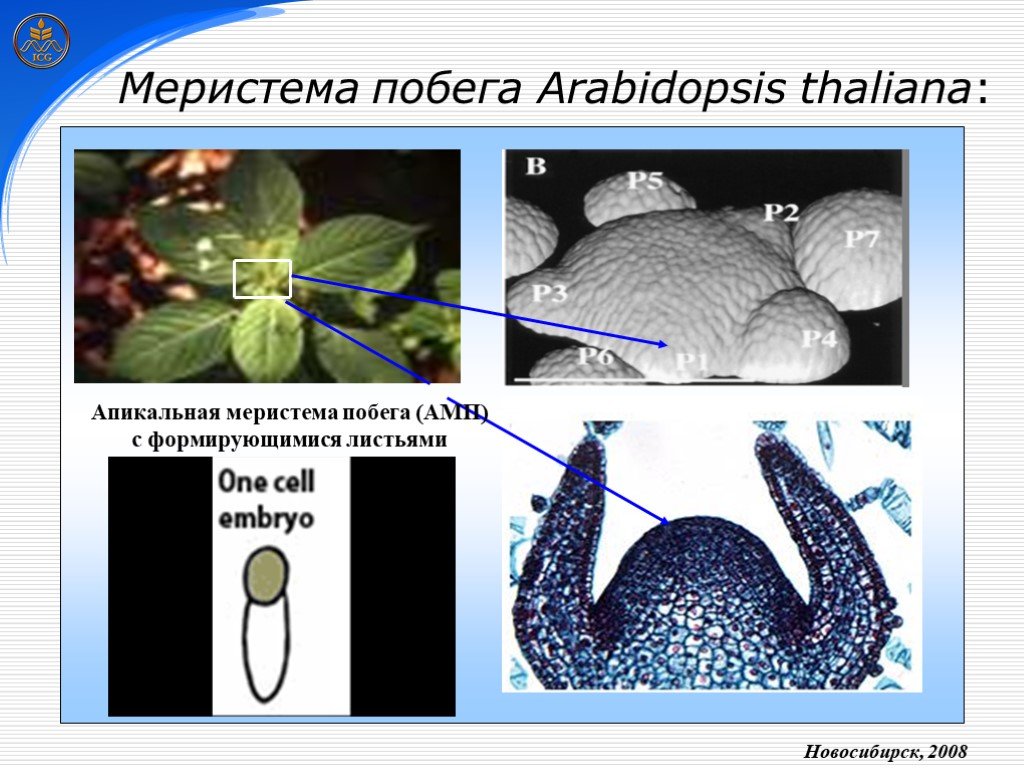 Слайд 3
Слайд 3 Слайд 4
Слайд 4 Слайд 5
Слайд 5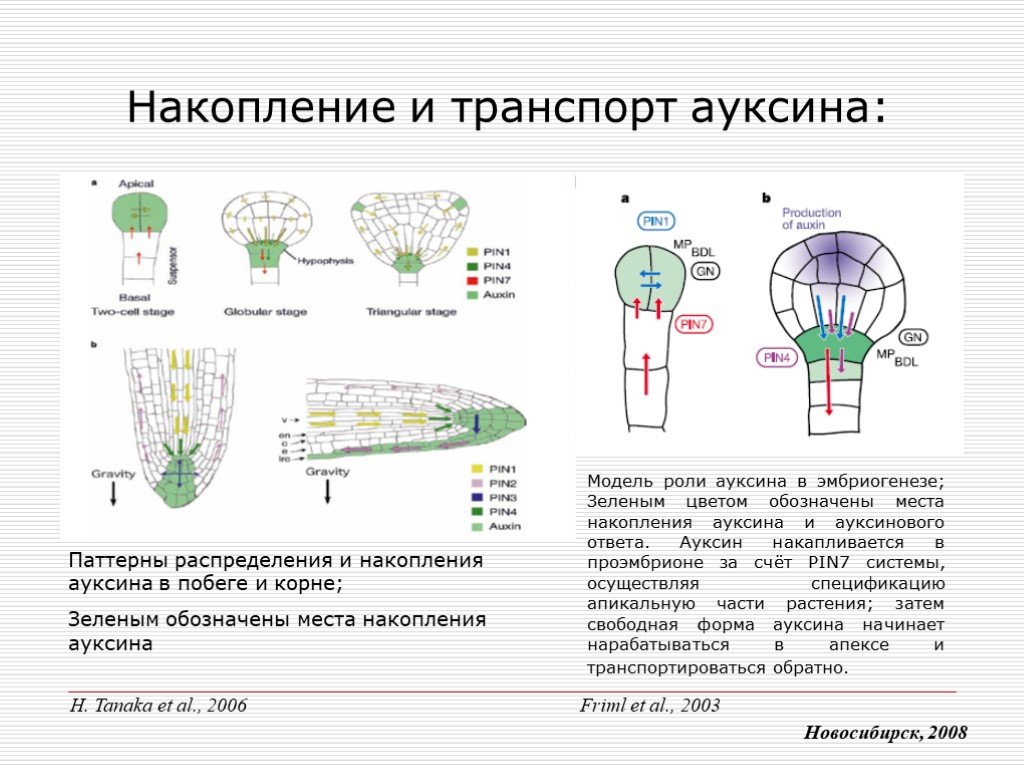 Слайд 6
Слайд 6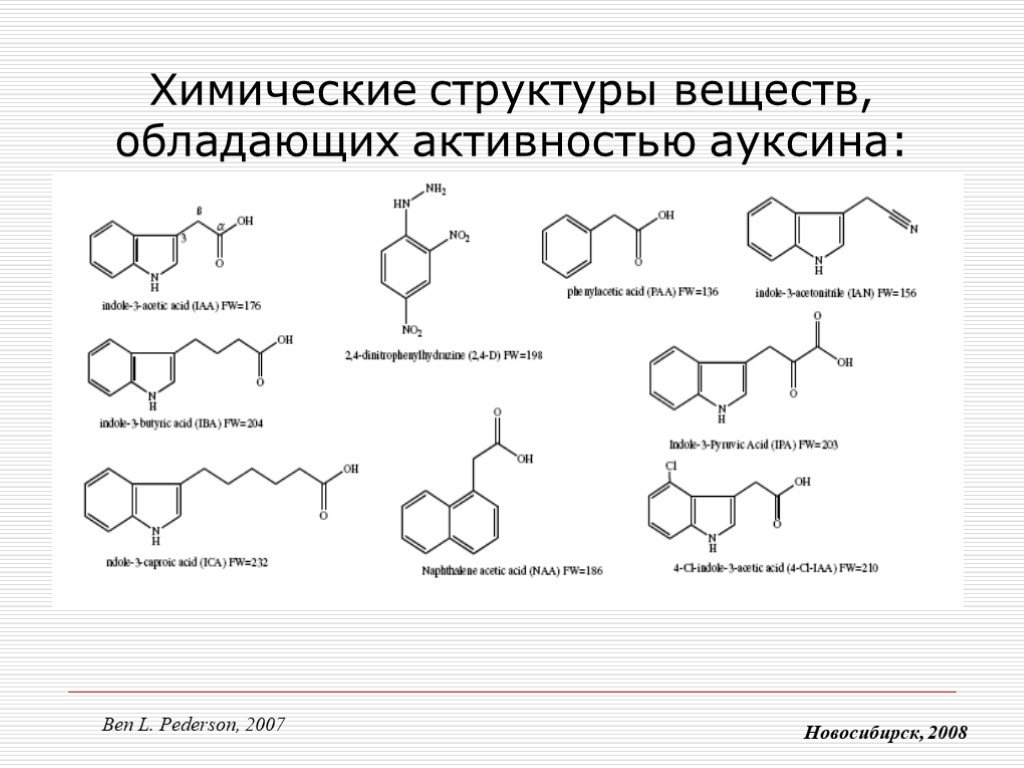 Слайд 7
Слайд 7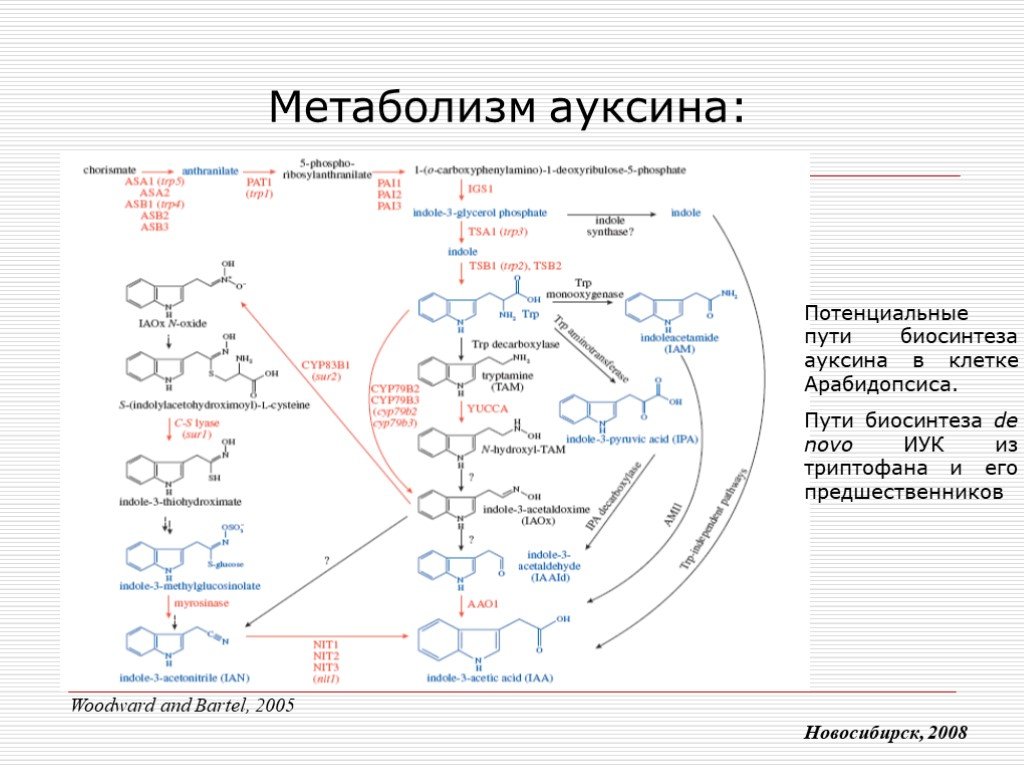 Слайд 8
Слайд 8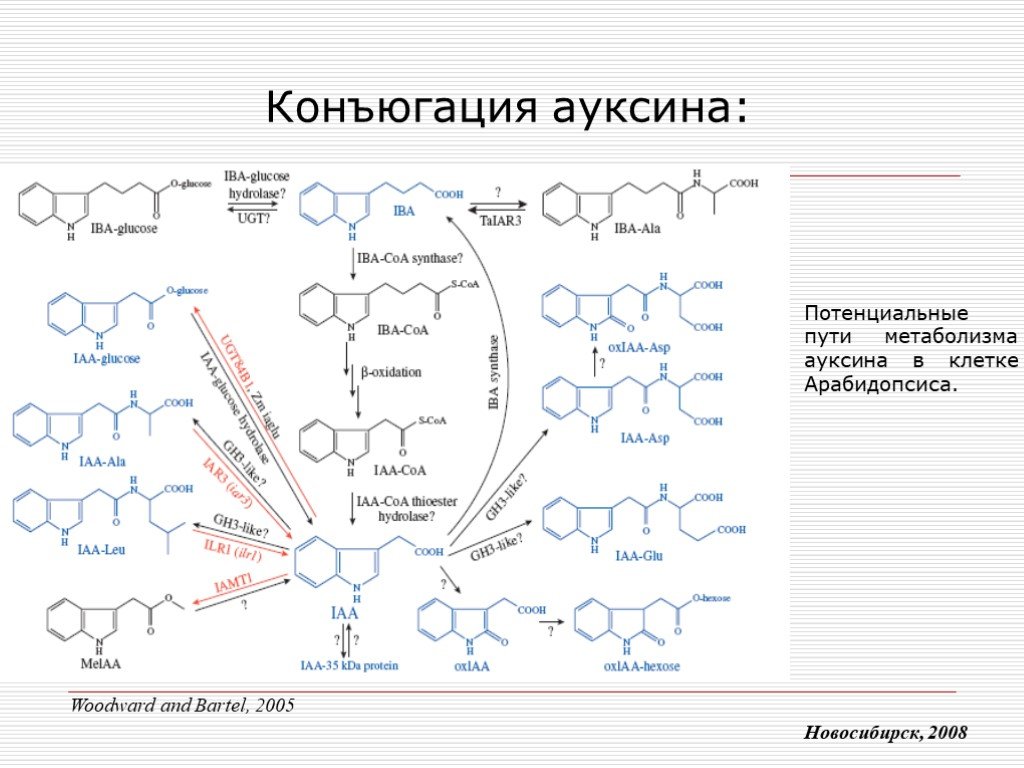 Слайд 9
Слайд 9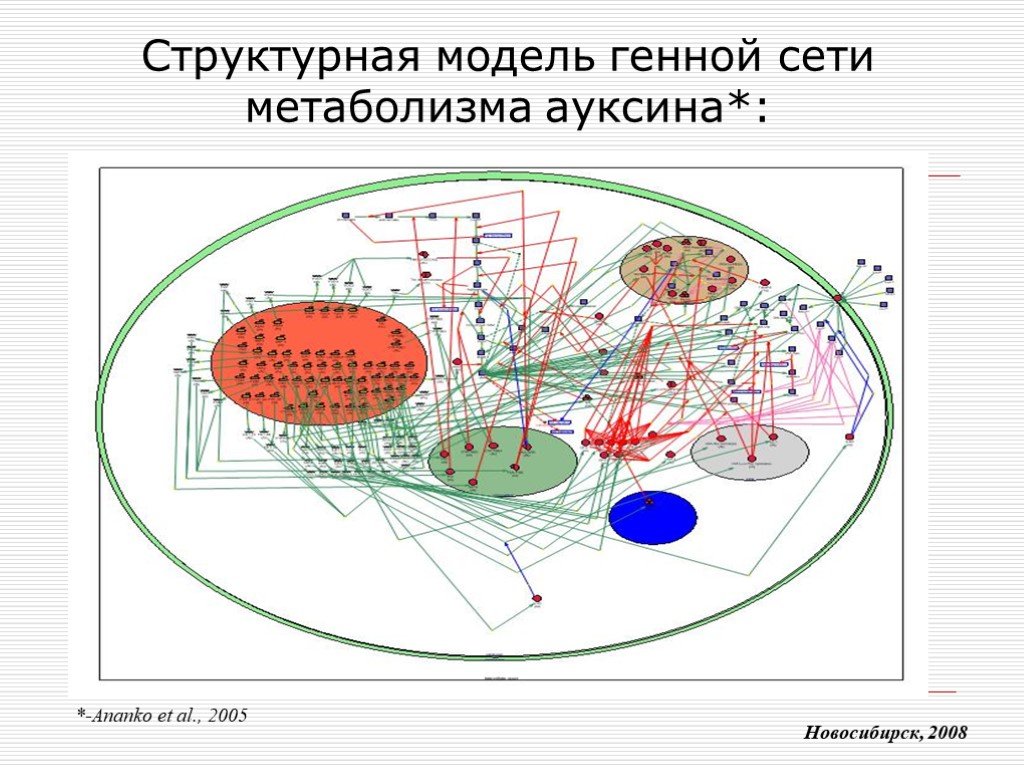 Слайд 10
Слайд 10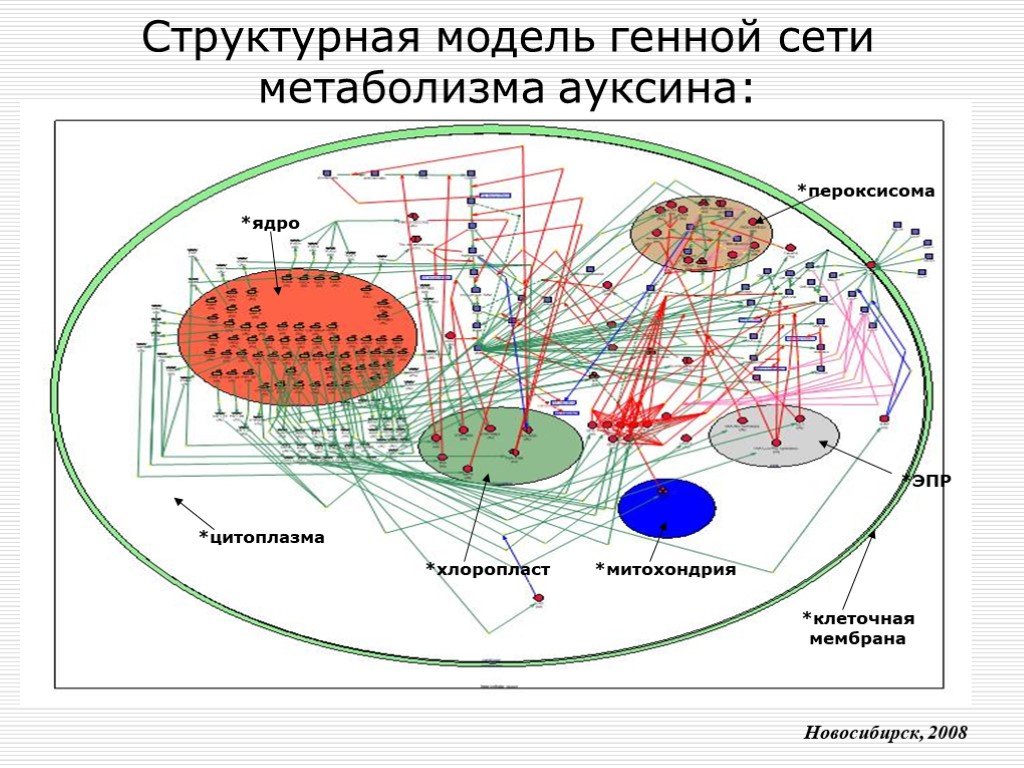 Слайд 11
Слайд 11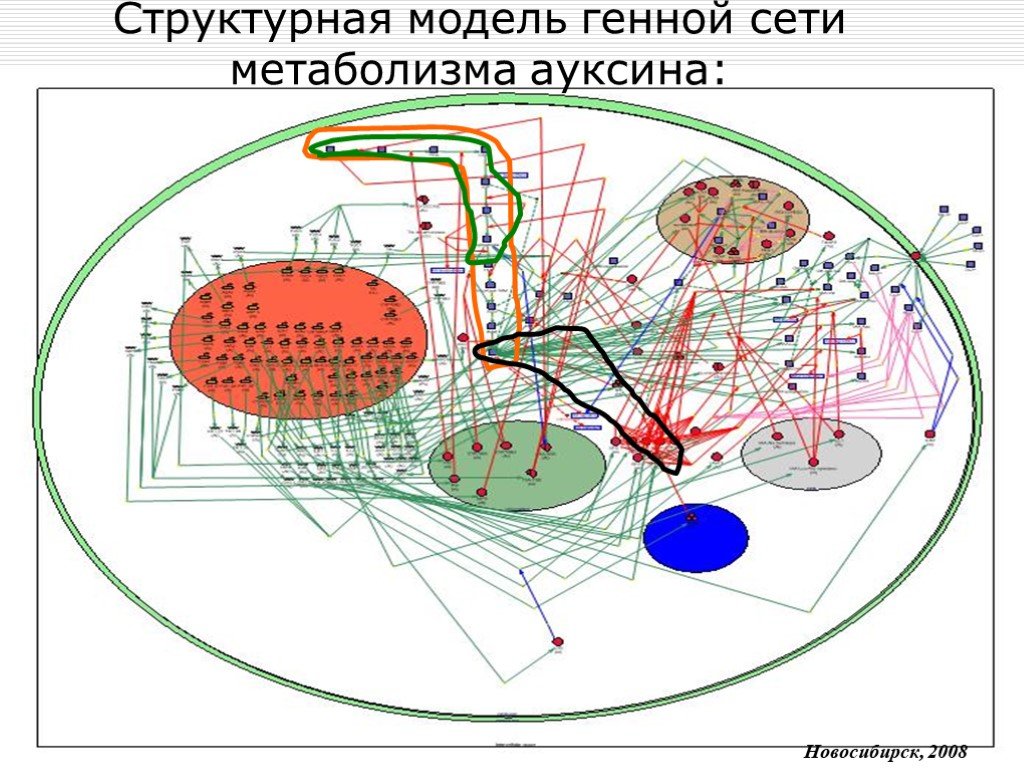 Слайд 12
Слайд 12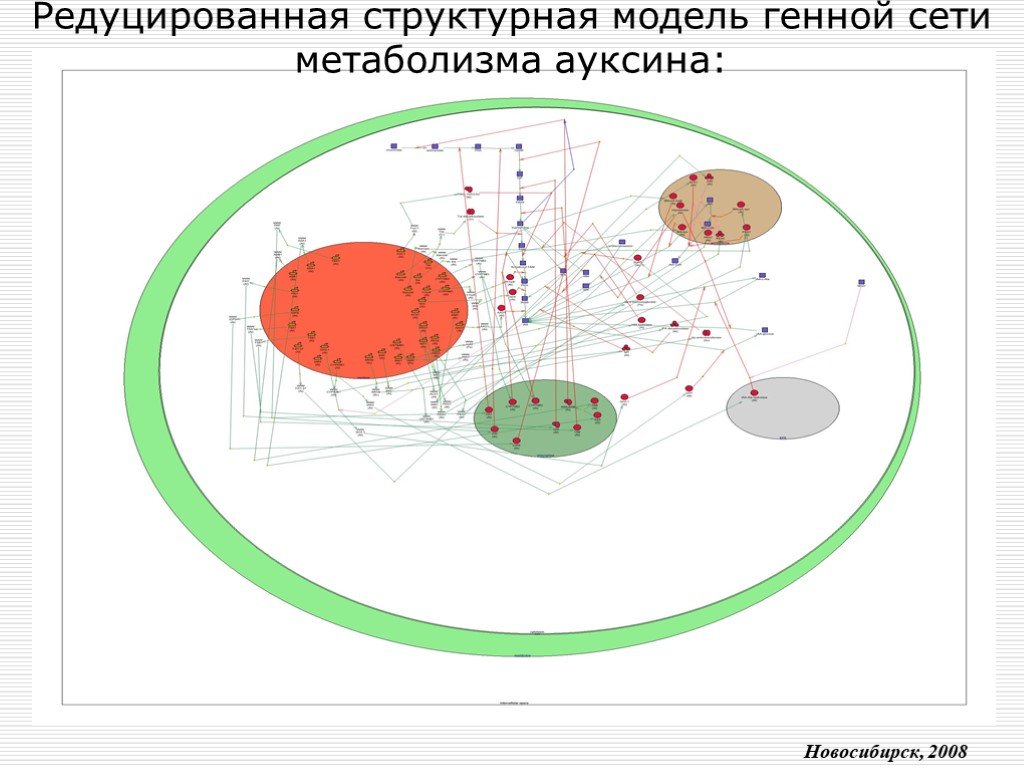 Слайд 13
Слайд 13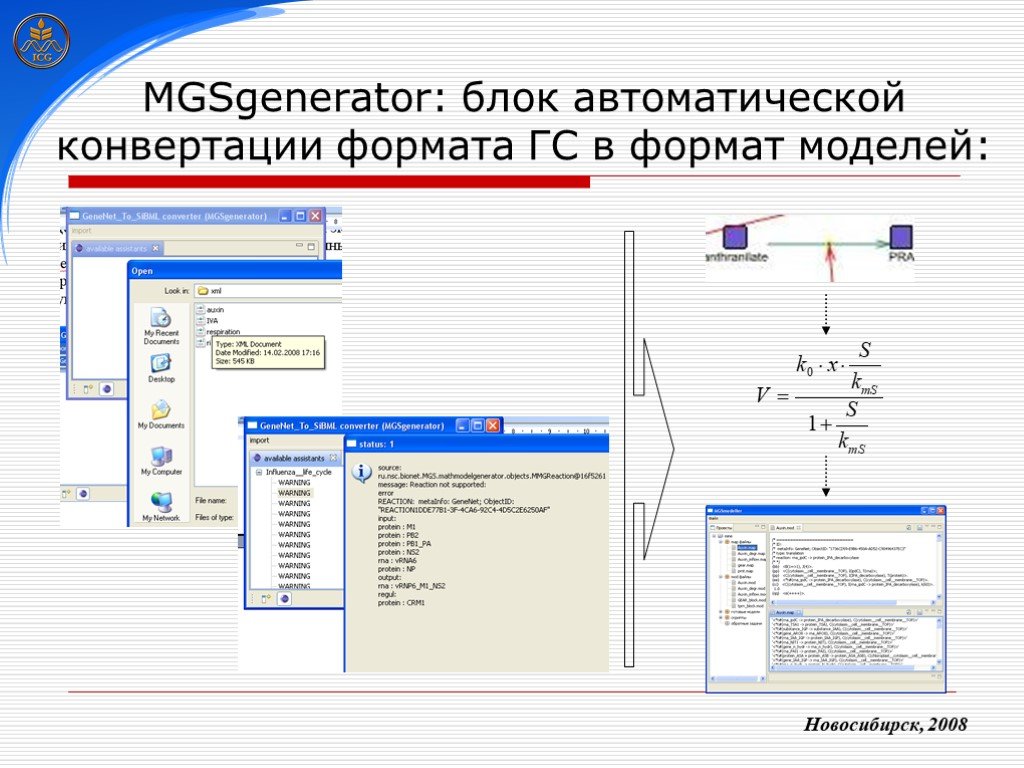 Слайд 14
Слайд 14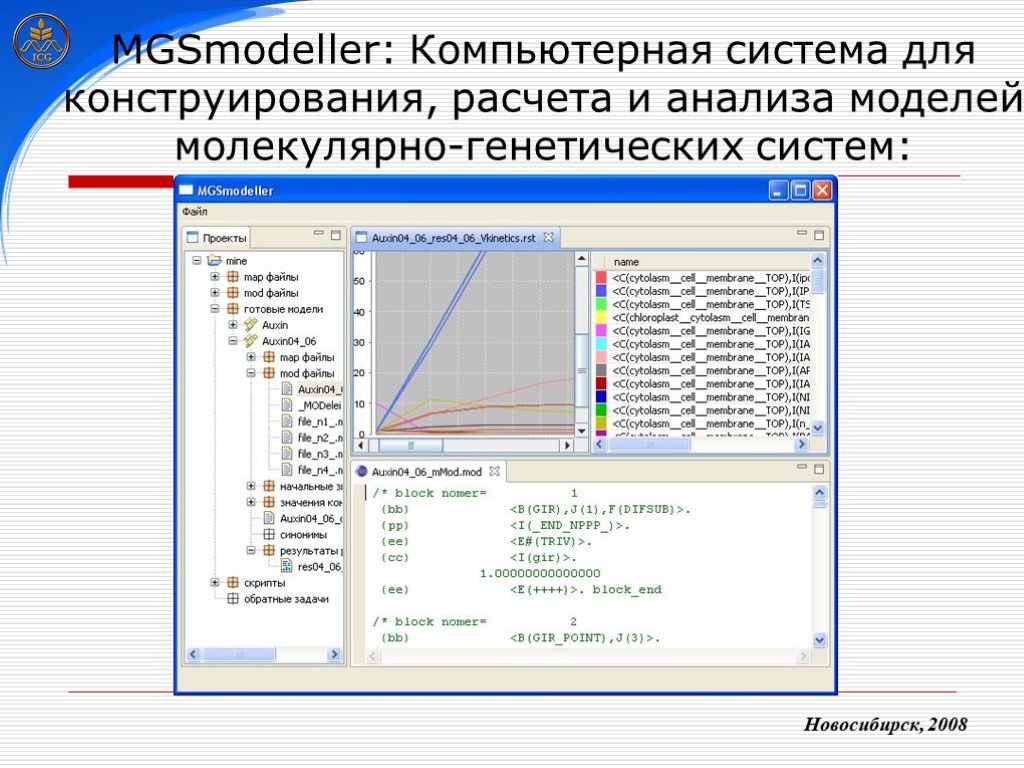 Слайд 15
Слайд 15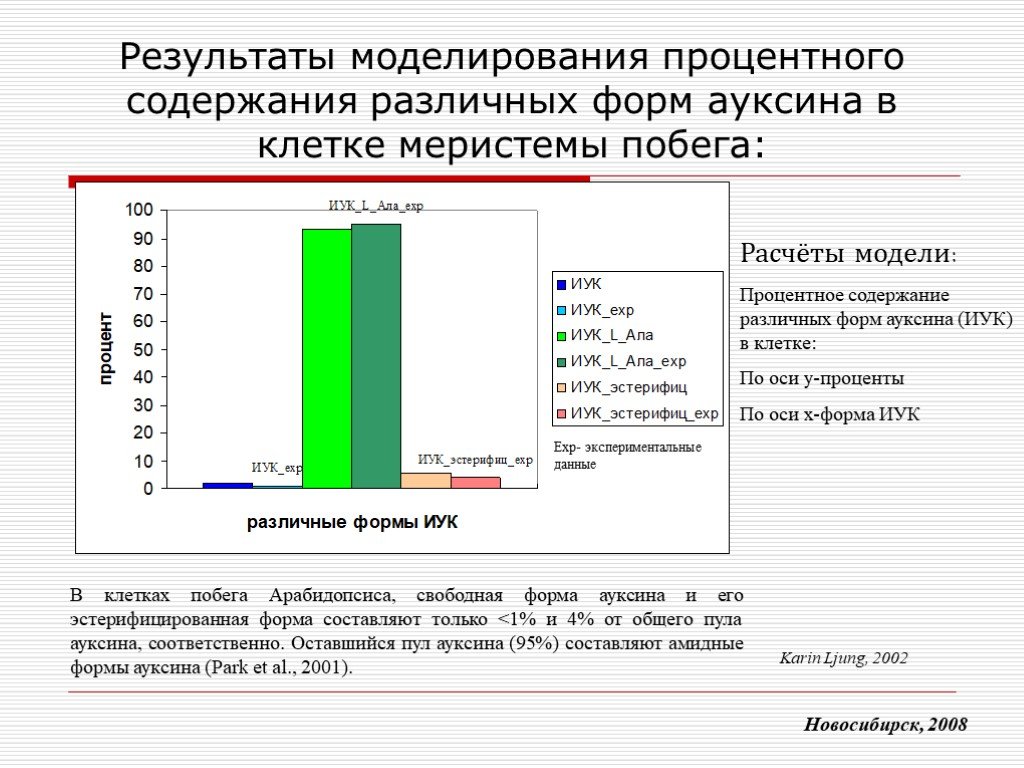 Слайд 16
Слайд 16 Слайд 17
Слайд 17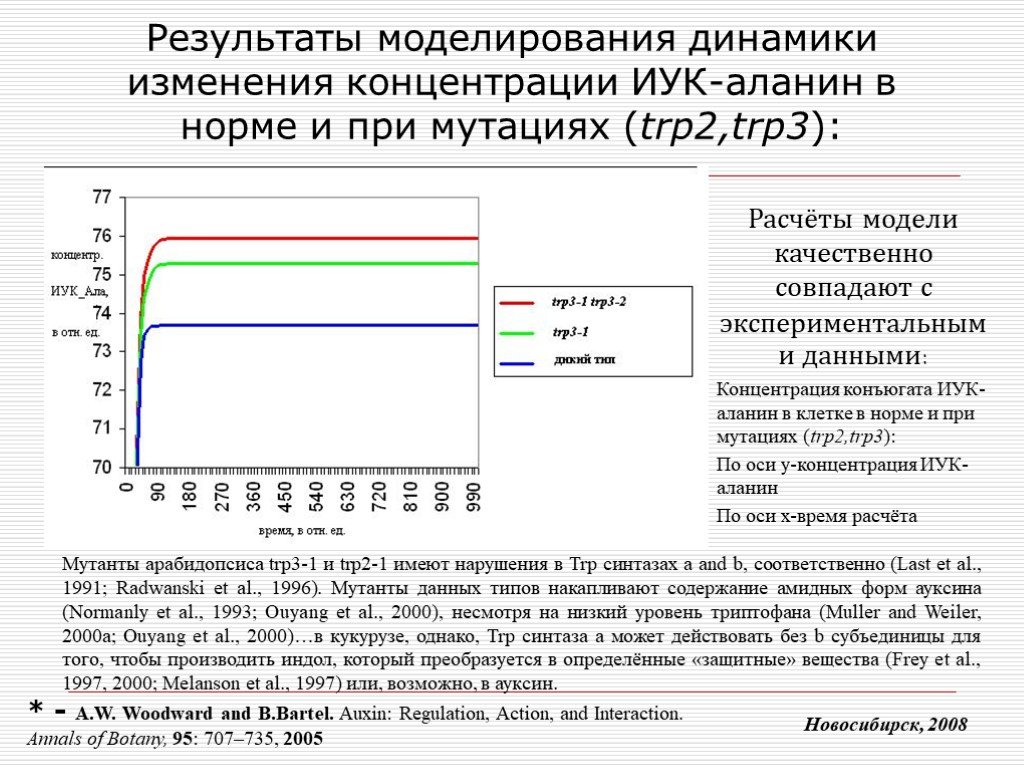 Слайд 18
Слайд 18 Слайд 19
Слайд 19 Слайд 20
Слайд 20 Слайд 21
Слайд 21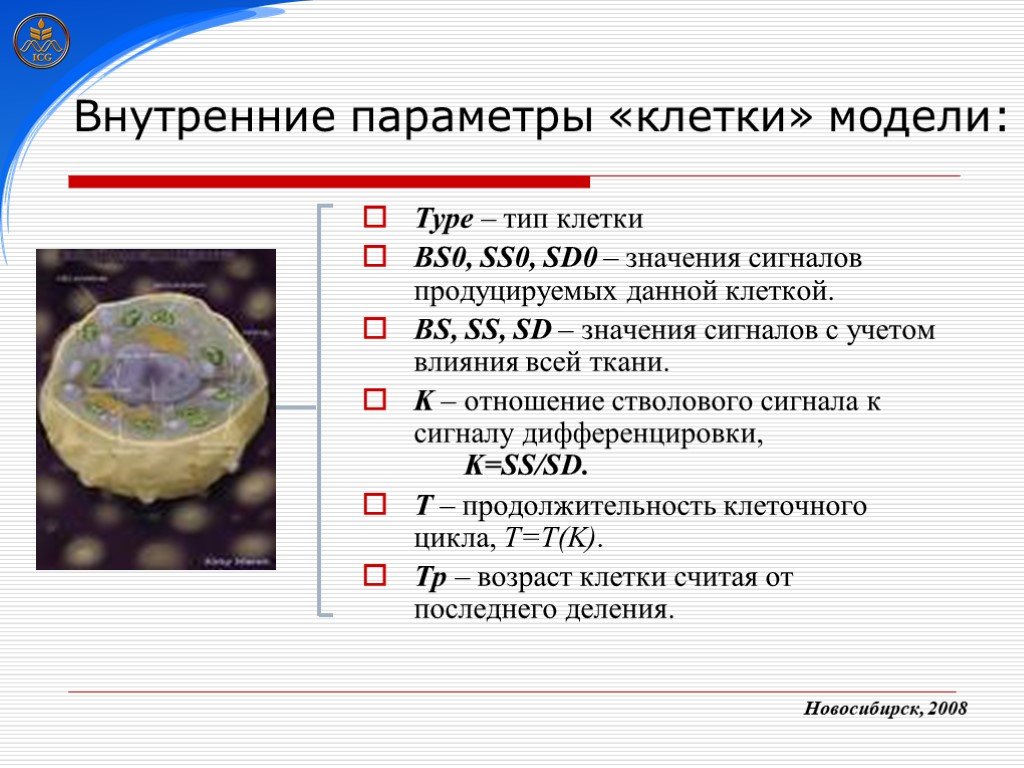 Слайд 22
Слайд 22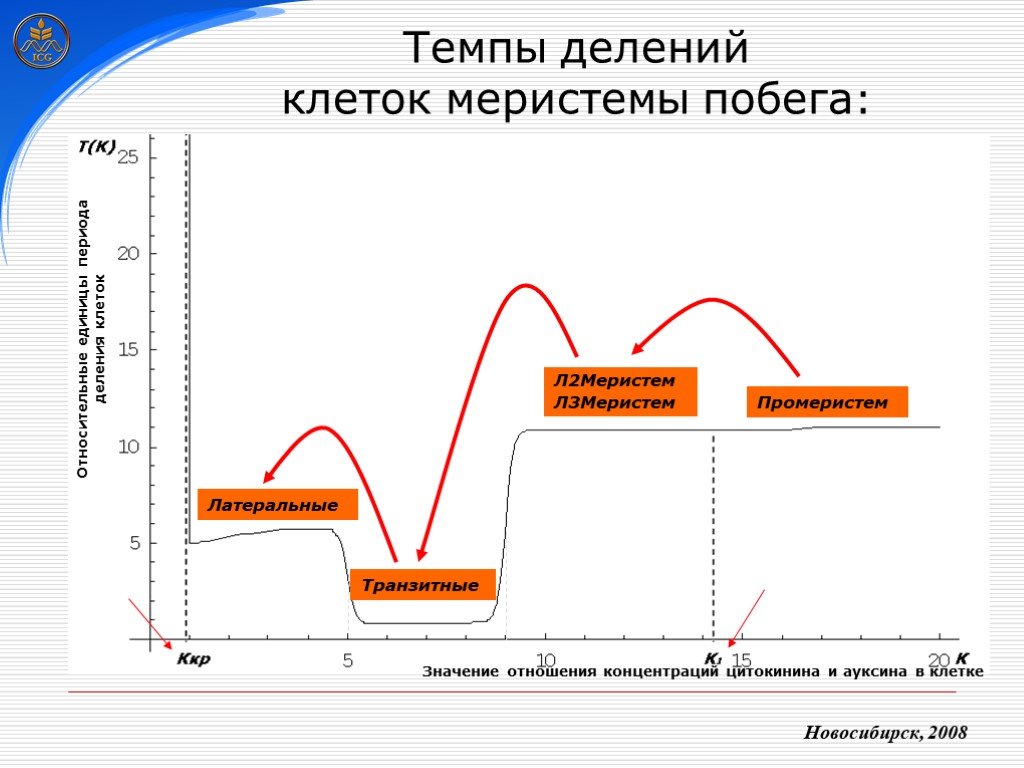 Слайд 23
Слайд 23 Слайд 24
Слайд 24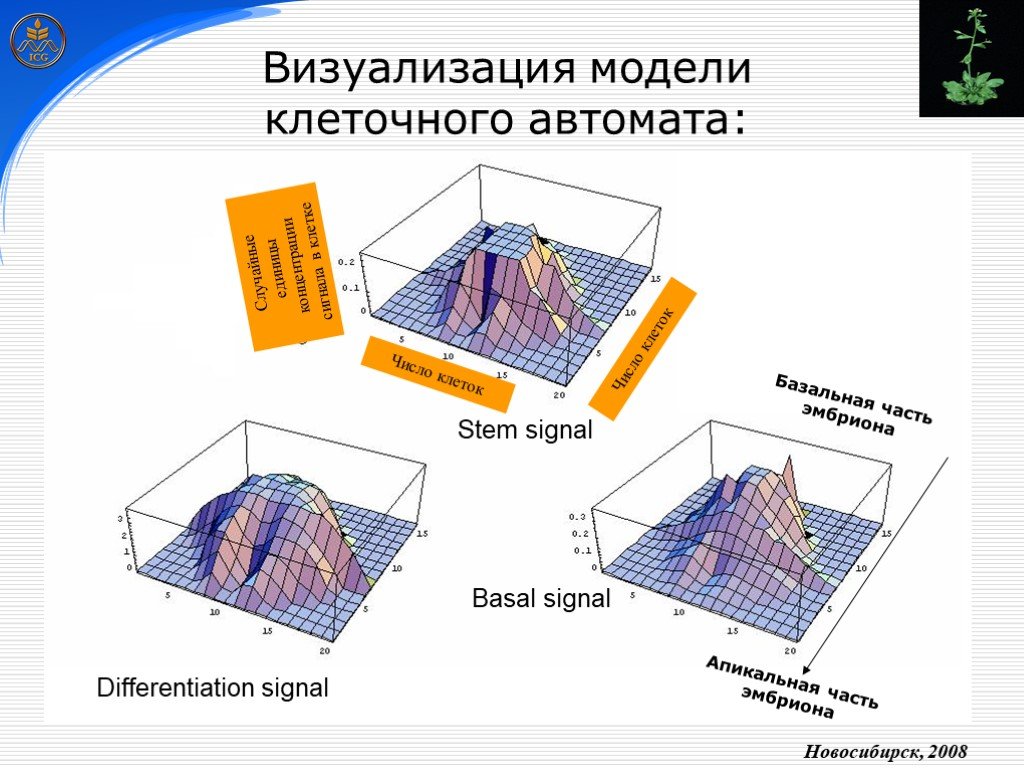 Слайд 25
Слайд 25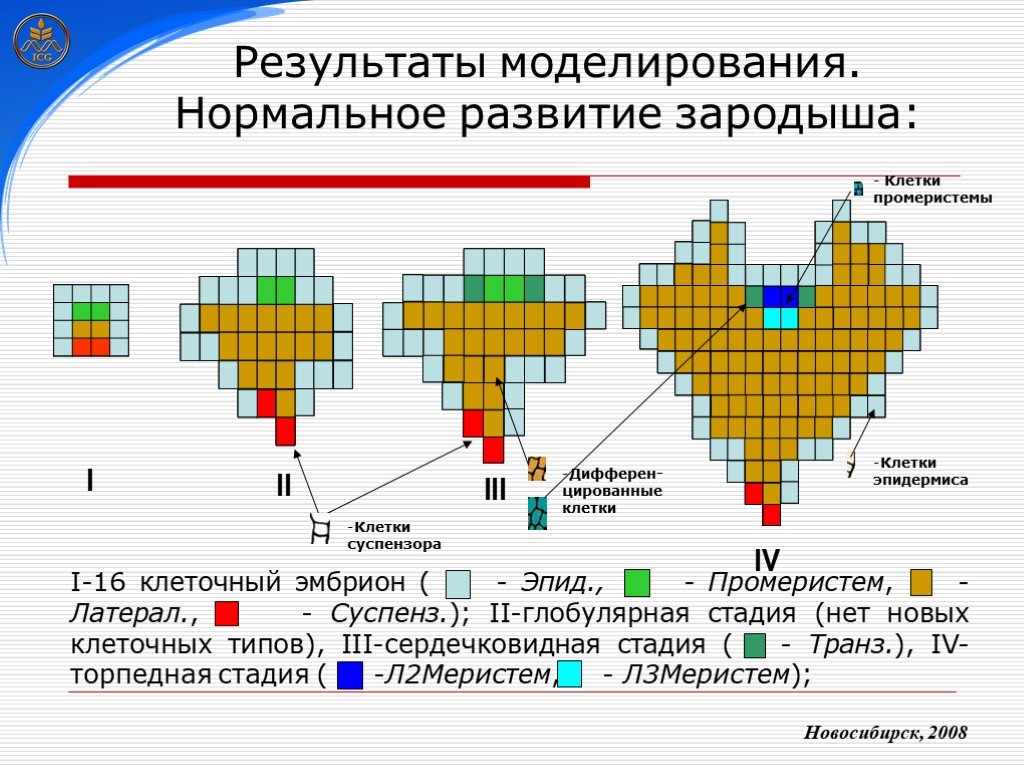 Слайд 26
Слайд 26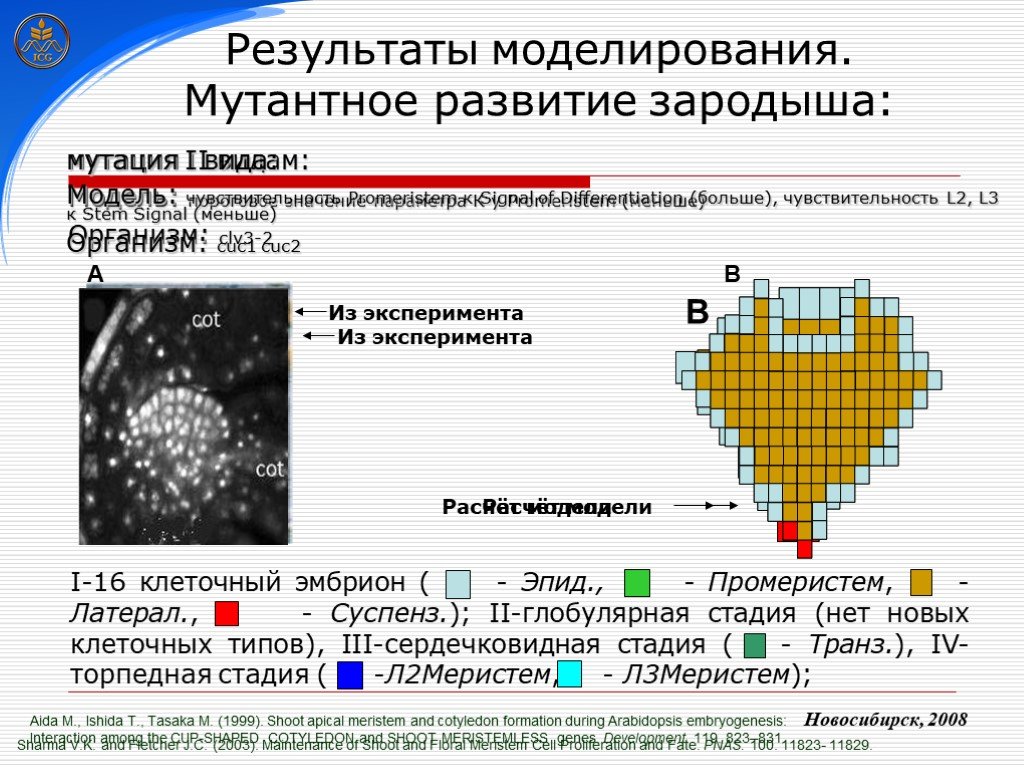 Слайд 27
Слайд 27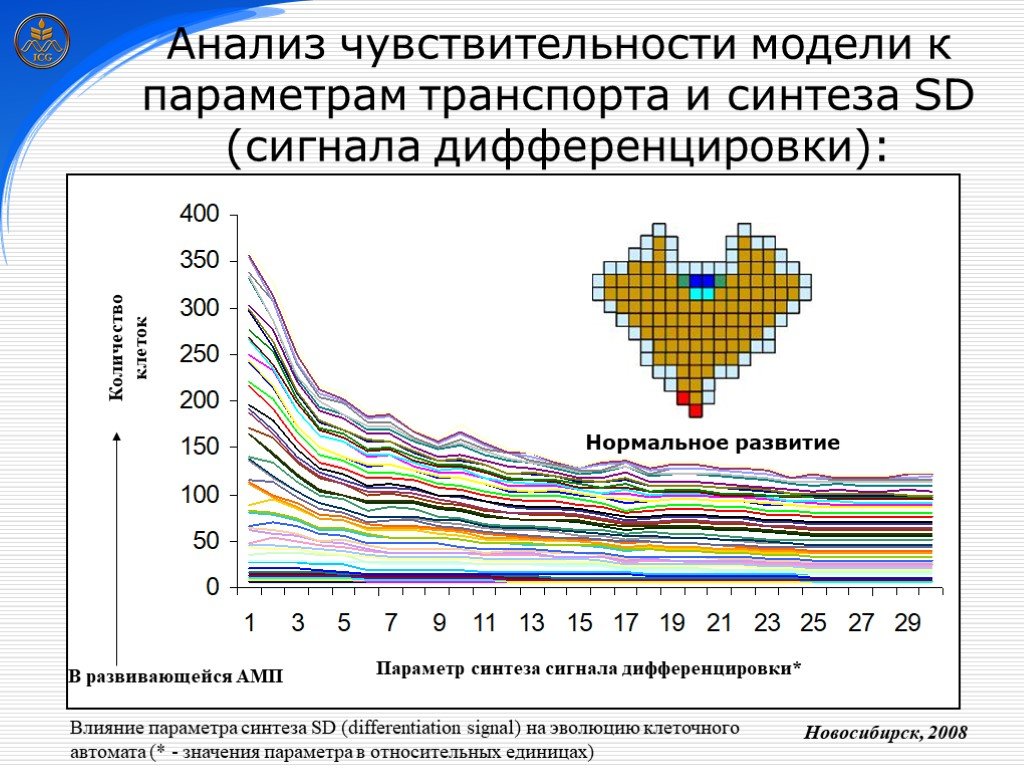 Слайд 28
Слайд 28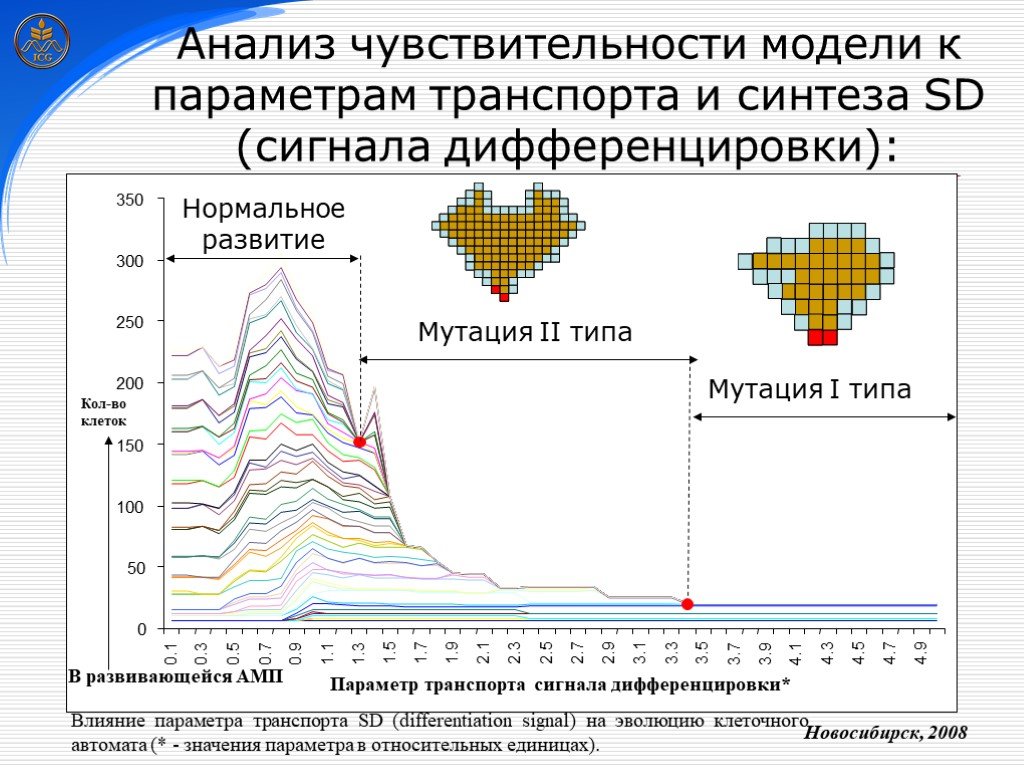 Слайд 29
Слайд 29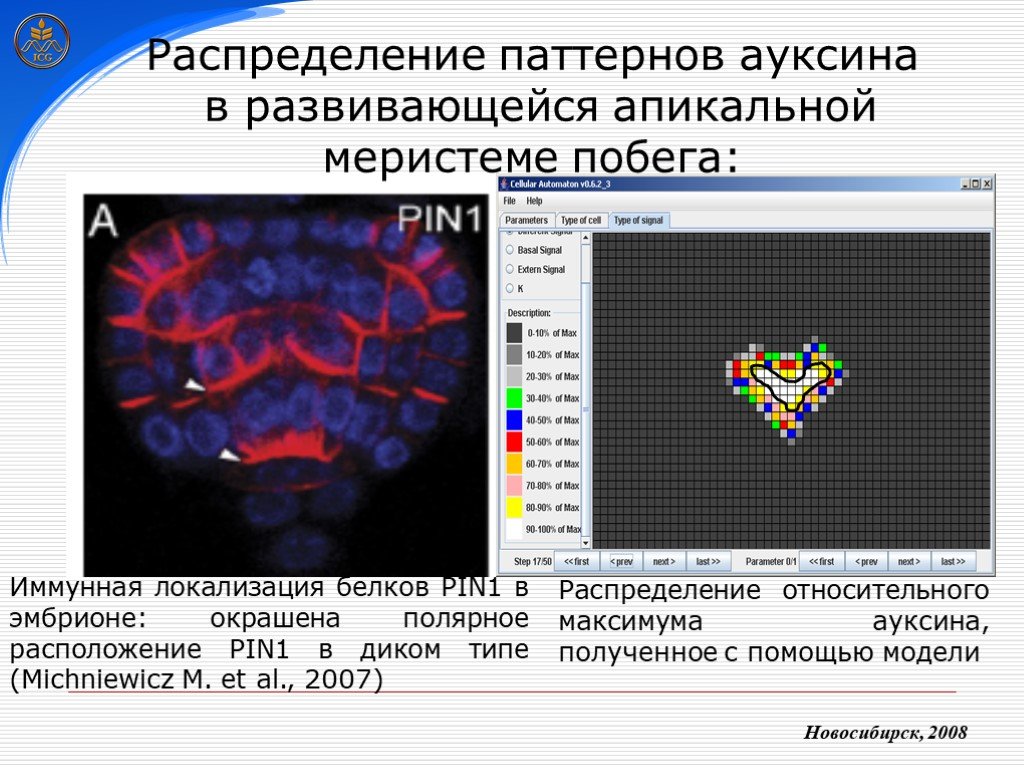 Слайд 30
Слайд 30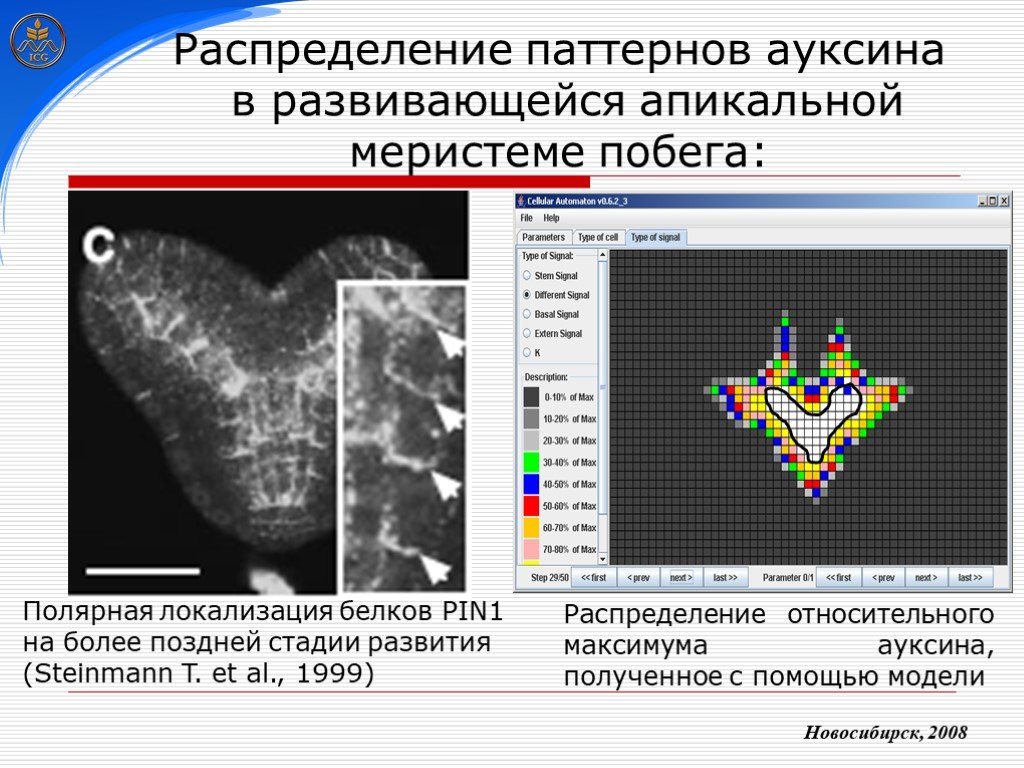 Слайд 31
Слайд 31 Слайд 32
Слайд 32 Слайд 33
Слайд 33 Слайд 34
Слайд 34 Слайд 35
Слайд 35Презентацию на тему "Моделирование регуляции развития меристемы побега" (9 класс) можно скачать абсолютно бесплатно на нашем сайте. Предмет проекта: Биология. Красочные слайды и иллюстрации помогут вам заинтересовать своих одноклассников или аудиторию. Для просмотра содержимого воспользуйтесь плеером, или если вы хотите скачать доклад - нажмите на соответствующий текст под плеером. Презентация содержит 35 слайд(ов).
Слайды презентации
Список похожих презентаций

Моделирование как средство саморазвития учащихся
« Единственный путь, ведущий к знанию – это деятельность» Б. Шоу. Формирование познавательной активности. Содержание учебного материала. Организация ...
Экологические проекты в программе развития моногородов
Байкальский ЦБК является знаковым проектом для оценки отношения России к вопросам экологии. официально обнаружены токсичные вещества, включённые в ...
Биология развития как функция времени
Биология развития — не что иное, как изучение каждой молекулы в клетке, органа и организма как функций времени. Теперь даже эволюцию рассматривают ...
Размножение и типы развития насекомых
тт Тля. Размножение. Бесполое и половое размножение. Половой диморфизм – сильное отличие самцов от самок. Откладка яиц насекомыми. Непрямое развитие ...
Типы развития насекомых
Выберите из предложенных изображений беспозвоночных животных насекомых. Какие части насекомого изображены? Вопросы урока. Как называется способ размножения, ...
Оценка пищевого статуса развития
Пищевой статус человека - это такое состояние его структуры, функции и адаптационных резервов, которые сложились под влиянием предшествующего фактического ...
Постэмбриональный период развития
Закрепить знания об эмбриональном развитии организма. Расширить знание учящихся о закономерностях послезародышевого развития огранизмов. Развивать ...
История возникновения и развития жизни на земле
ИСТОРИЯ ВОЗНИКНОВЕНИЯ И РАЗВИТИЯ ЖИЗНИ НА ЗЕМЛЕ. Когда примерно образовалась наша планета? Как вы считаете она была горячей или холодной? За счет ...
История и основные этапы развития возрастной физиологии
Научное изучение возрастных особенностей детского организма началось сравнительно недавно – во второй половине XIX в. Вскоре после открытия закона ...
Зависимость роста и развития растений от условий окружающей среды
Стихотворение посвящённое временам года. Всё начинается с весны С капелей ледяного снега И первый маленький росток Пробьётся сквозь сугробы снега. ...
Идея развития органического мира в биологии
Эволюционное учение. - Это наука о причинах, движущих силах и общих закономерностях исторического развития живой природы. Чарлз Роберт Дарвин (1809-1882). ...
Генетика. история развития генетики. основные понятия
Грегор Мендель (1822 - 1884г.г.). МАОУ лицей №8 им.Н.Н. Рукавишникова г. Томска, Батракова К.А. Г. де Фриз К. Корренс Э. Чермак. 1906г. – У. Бетсон ...
Генетика: история развития науки. Основные понятия
ЗАДАЧИ УРОКА:. Познакомиться с наукой «генетика», ее историей и достижениями. Определить цели и задачи генетики в современном мире. Показать роль ...
Биотехнология, её достижения и перспектива развития
Биотехнология — это производство необходимых человеку продуктов и материалов с помощью живых организмов, культивируемых клеток и биологических процессов. ...
Нарушения нейрогуморальной регуляции
Нарушения функций желез внутренней секреции сопровождаются уменьшением при гипофункции или увеличением при гиперфункции производства гормонов. Уменьшение ...
История развития анатомии, физиологии и медицины
(около 460 до н. э., остров Кос — около 377 до н. э.) древнегреческий врач, «отец медицины», которая выделилась из философии в отдельную науку. Гиппократ. ...
Основные этапы развития животного мира
§ 60. Доказательства эволюции животного мира. Учение Ч. Дарвина об эволюции. 1. Расскажите об основных стадиях эмбрионального развития позвоночных, ...
История развития биологии
Цель урока:. Познакомиться с развитием биологии как науки, с глубокой древности и до наших дней, на основе биологических исследований и открытий великих ...
Побег.строение и значение побега
Побег – это стебель с расположенными на нем листьями и почками. Строение побега. Узел – участок стебля, от которого отходит лист. Междоузлие – участок ...
История развития биологии
Цель урока:. 1.Познакомиться с задачами и содержанием курса «Основы общей биологии». 2. Осознать понимание биологических исследований для человека ...Конспекты
Роль эндокринной регуляции
6. . . Тема урока : «Роль эндокринной регуляции». Цели урока:. . - обеспечить формирование у учащихся знаний о роли желез внутренней секреции ...Происхождение и начальные этапы развития жизни на Земле ОПРЕДЕЛЕНИЕ ЖИЗНИ. ОСНОВНЫЕ СВОЙСТВА ЖИВЫХ ОРГАНИЗМОВ. УРОВНИ ОРГАНИЗАЦИИ ЖИЗНИ
Урок № 1. Тема урока: Глава 1. Происхождение и начальные этапы. развития жизни на Земле. ОПРЕДЕЛЕНИЕ ЖИЗНИ. ОСНОВНЫЕ СВОЙСТВА ЖИВЫХ ОРГАНИЗМОВ. ...Размножение насекомых. Типы развития насекомых
Урок по теме: "Размножение насекомых. Типы развития насекомых"7 класс. Задачи урока:. закрепление классификационных признаков представителей класса ...Постоянство температуры тела и способы ее регуляции
Смирнова Евгения Павловна, учитель биологии. ГБОУ гимназия №446 Колпинского района Санкт-Петербурга. Тема урока: «Постоянство температуры тела ...Предупреждение искривления позвоночника и развития плоскостопия
ГБС(К)ОУ школа –интернат № 7восьмого вида станицы Казанской. Краснодарского края. Конспект урока по биологии на тему "Предупреждение искривления. ...Условия развития познавательной активности школьников на уроках биологии
. РЕФЕРАТ. ТЕМА: Условия развития познавательной активности школьников на уроках биологии. Выполнил: Захаров ...Лист – часть побега, его внешнее и внутреннее строение
Урок по теме «Лист – часть побега, его внешнее и внутреннее строение». Цель:. . . сформировать понятия, связанные с морфологией листа, познакомить ...Особенности строения, цикл развития и распространение зеленого мха
Открытый урок. Тема:. «Особенности строения, цикл развития и распространение зеленого мха». . Цель урока:. . сформировать представление ...Первоначальные и современные представления о происхождении жизни. Начальные этапы развития жизни
Конспект, рассчитанный на 2 урока по биологии в 9 классе. Жилевская Инна Владимировна. ,. . учитель биологии. . первой категории. КГУ «Средняя ...Начальные этапы развития жизни
Муниципальное бюджетное общеобразовательное учреждение. « Средняя общеобразовательная школа № 93». города Новокузнецка Кемеровской области. ...Советы как сделать хороший доклад презентации или проекта
- Постарайтесь вовлечь аудиторию в рассказ, настройте взаимодействие с аудиторией с помощью наводящих вопросов, игровой части, не бойтесь пошутить и искренне улыбнуться (где это уместно).
- Старайтесь объяснять слайд своими словами, добавлять дополнительные интересные факты, не нужно просто читать информацию со слайдов, ее аудитория может прочитать и сама.
- Не нужно перегружать слайды Вашего проекта текстовыми блоками, больше иллюстраций и минимум текста позволят лучше донести информацию и привлечь внимание. На слайде должна быть только ключевая информация, остальное лучше рассказать слушателям устно.
- Текст должен быть хорошо читаемым, иначе аудитория не сможет увидеть подаваемую информацию, будет сильно отвлекаться от рассказа, пытаясь хоть что-то разобрать, или вовсе утратит весь интерес. Для этого нужно правильно подобрать шрифт, учитывая, где и как будет происходить трансляция презентации, а также правильно подобрать сочетание фона и текста.
- Важно провести репетицию Вашего доклада, продумать, как Вы поздороваетесь с аудиторией, что скажете первым, как закончите презентацию. Все приходит с опытом.
- Правильно подберите наряд, т.к. одежда докладчика также играет большую роль в восприятии его выступления.
- Старайтесь говорить уверенно, плавно и связно.
- Старайтесь получить удовольствие от выступления, тогда Вы сможете быть более непринужденным и будете меньше волноваться.
Информация о презентации
Дата добавления:18 сентября 2019
Категория:Биология
Классы:
Содержит:35 слайд(ов)
Поделись с друзьями:
Скачать презентацию









