Презентация "Популяционно-статистический метод" по биологии – проект, доклад
 Слайд 1
Слайд 1 Слайд 2
Слайд 2 Слайд 3
Слайд 3 Слайд 4
Слайд 4 Слайд 5
Слайд 5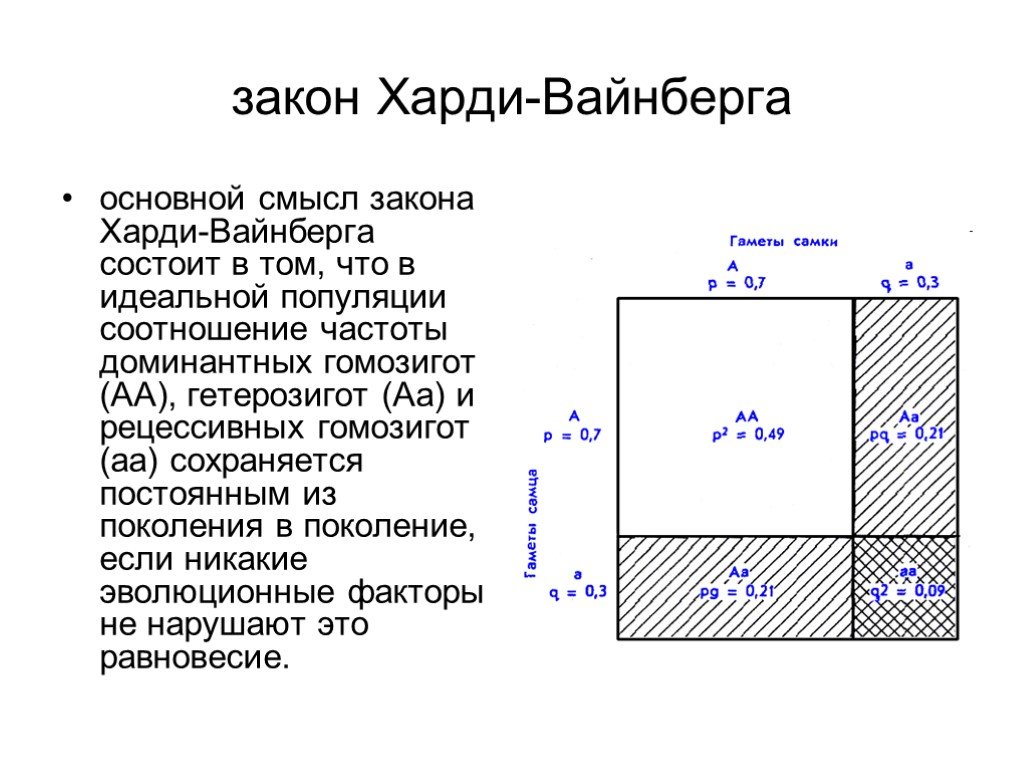 Слайд 6
Слайд 6 Слайд 7
Слайд 7 Слайд 8
Слайд 8 Слайд 9
Слайд 9 Слайд 10
Слайд 10 Слайд 11
Слайд 11 Слайд 12
Слайд 12 Слайд 13
Слайд 13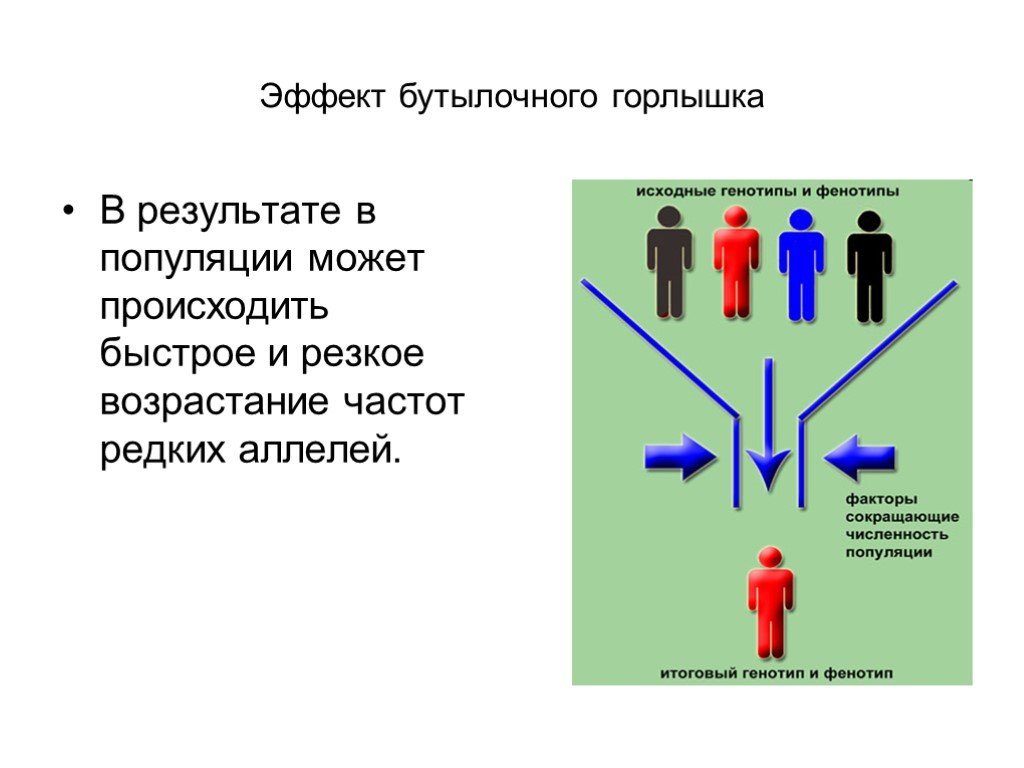 Слайд 14
Слайд 14 Слайд 15
Слайд 15 Слайд 16
Слайд 16 Слайд 17
Слайд 17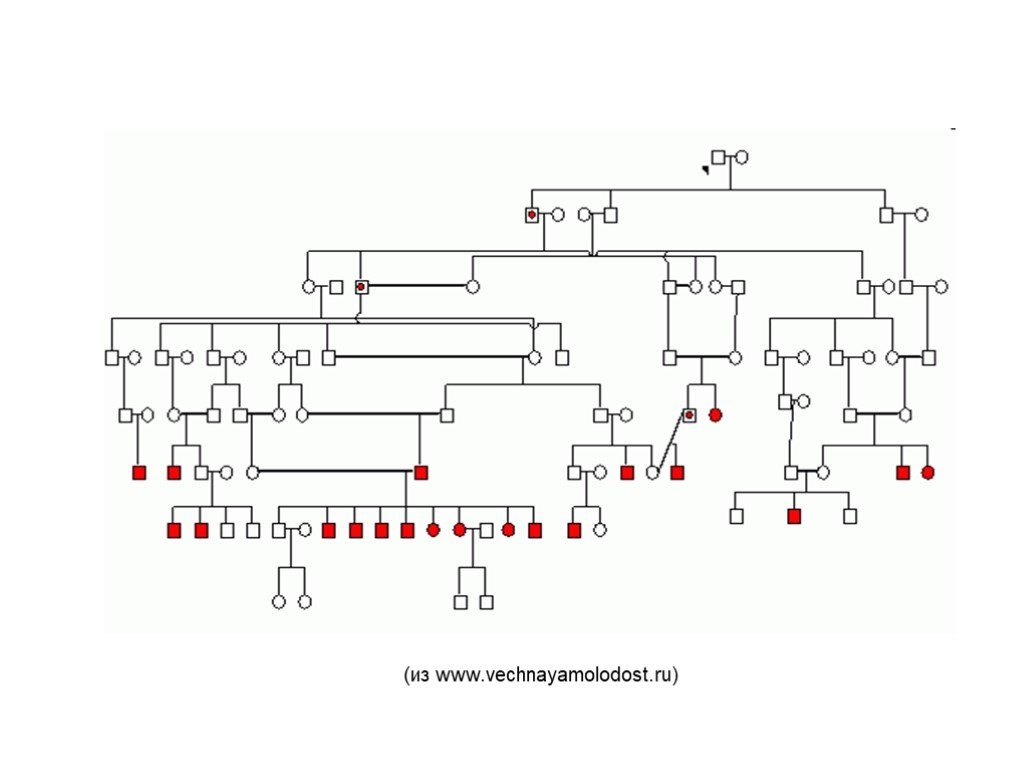 Слайд 18
Слайд 18 Слайд 19
Слайд 19 Слайд 20
Слайд 20 Слайд 21
Слайд 21 Слайд 22
Слайд 22 Слайд 23
Слайд 23 Слайд 24
Слайд 24 Слайд 25
Слайд 25 Слайд 26
Слайд 26 Слайд 27
Слайд 27 Слайд 28
Слайд 28 Слайд 29
Слайд 29 Слайд 30
Слайд 30 Слайд 31
Слайд 31 Слайд 32
Слайд 32 Слайд 33
Слайд 33 Слайд 34
Слайд 34 Слайд 35
Слайд 35 Слайд 36
Слайд 36 Слайд 37
Слайд 37 Слайд 38
Слайд 38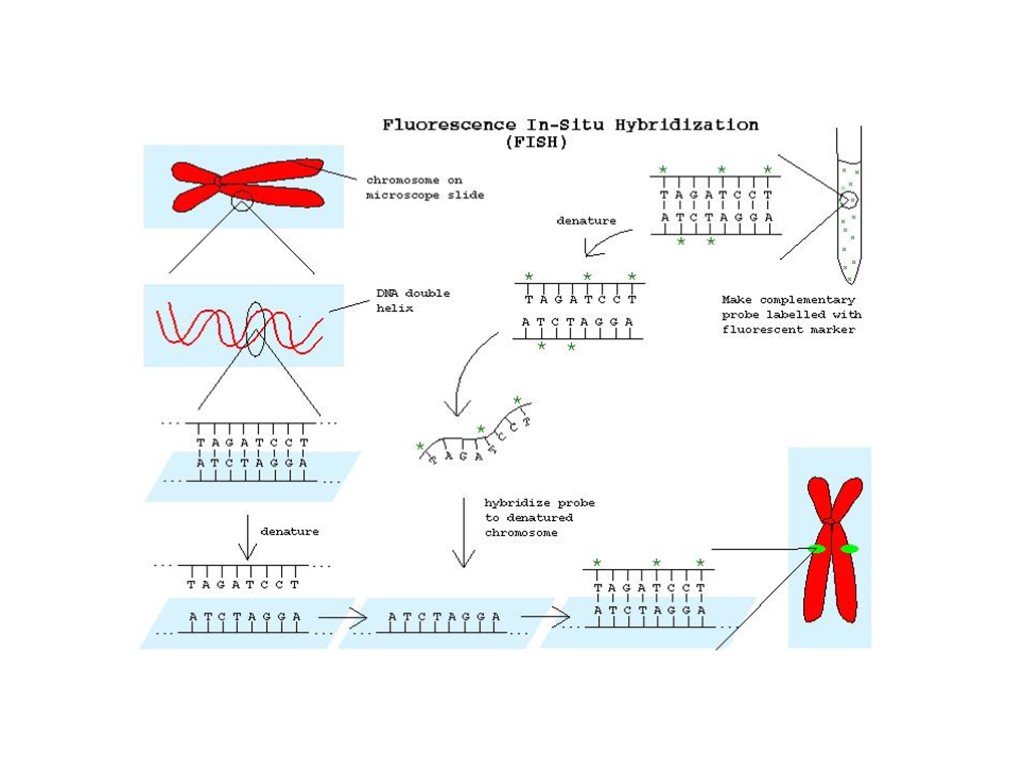 Слайд 39
Слайд 39 Слайд 40
Слайд 40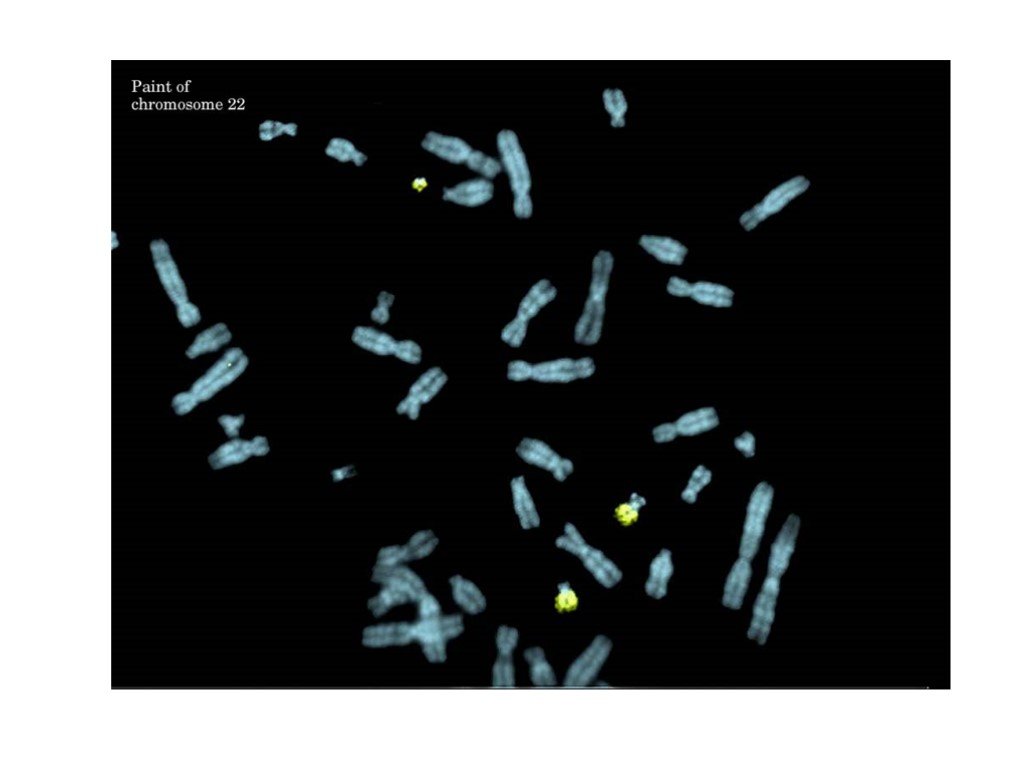 Слайд 41
Слайд 41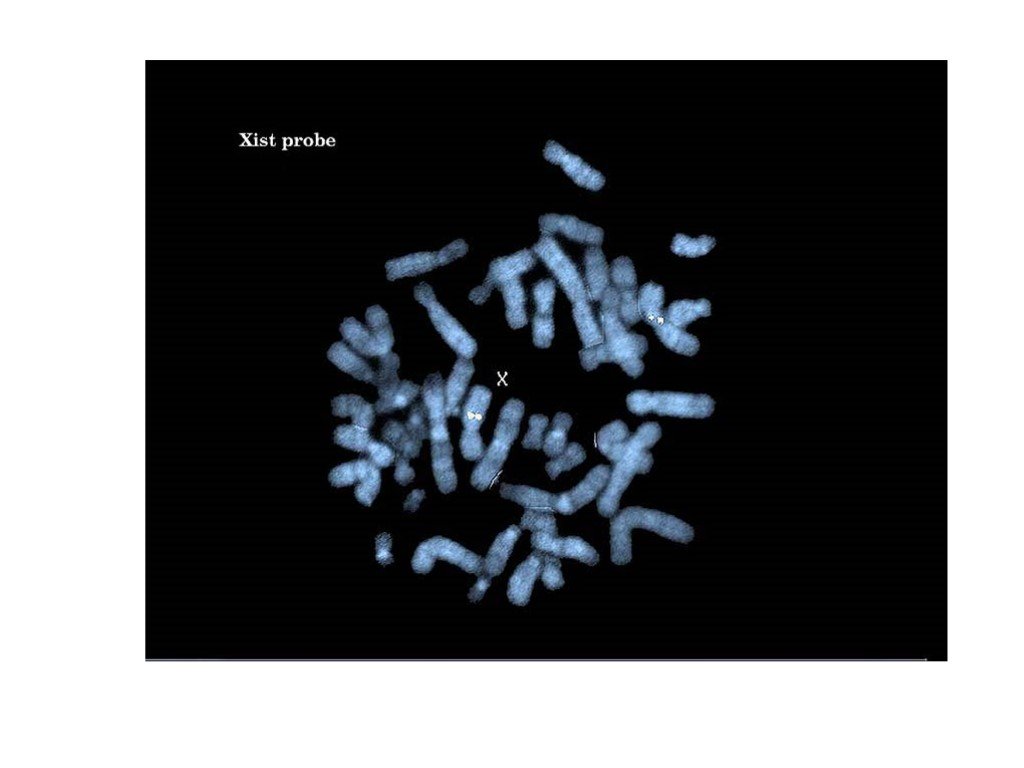 Слайд 42
Слайд 42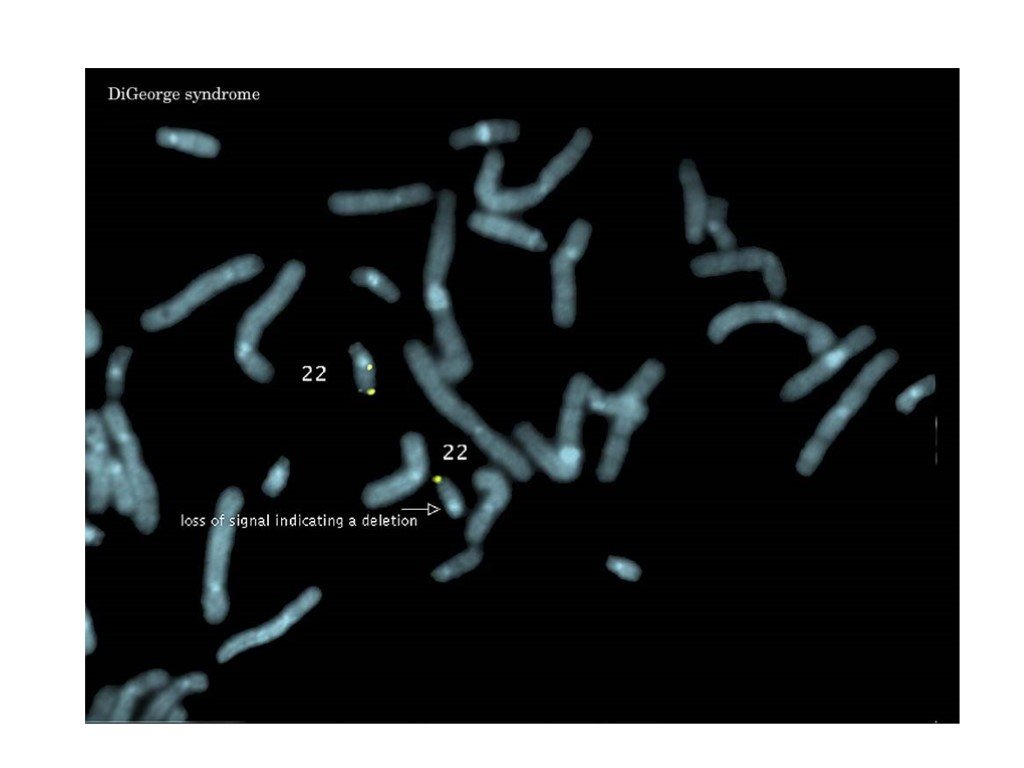 Слайд 43
Слайд 43Презентацию на тему "Популяционно-статистический метод" можно скачать абсолютно бесплатно на нашем сайте. Предмет проекта: Биология. Красочные слайды и иллюстрации помогут вам заинтересовать своих одноклассников или аудиторию. Для просмотра содержимого воспользуйтесь плеером, или если вы хотите скачать доклад - нажмите на соответствующий текст под плеером. Презентация содержит 43 слайд(ов).
Слайды презентации
Список похожих презентаций

Генеалогический метод
Генеалогия в широком смысле слова - родословная. Генеалогический метод - метод родословных, т.е. прослеживание болезни (или признака) в семье или ...
Моделирование, как метод изучения биологии
Виды моделей. Натуральные (материальные). информационные. Модель – это система, которая является образом, подобием некой дугой, например, природной ...
Дельфинотерапия как нетрадиционный метод психотерапии
Доказать, что дельфинотерапия может ускорить процесс излечения детей с различными психическими травмами и заболеваниями. Цель:. ●изучить источники ...
Генеалогический метод исследования
Генеалогический метод - это изучение родословной человека. Генеалогический метод состоит в изучении родословных на основе Менделеевских законов наследования ...
Генеалогический метод в генетике человека
Цели и задачи исследования. Выявить особенности наследования различных признаков, используя родословные: Составить родословную своей семьи; Определить ...
Близнецовый метод
Близнецовый метод изучения наследственности человека состоит в изучении развития признаков у близнецов. Что в человеке является врожденным, а что ...
Молекулярная биология гена
План лекции:. Ген – определение, классификация. Понятие о мутоне, реконе, цистроне. Строение гена у про- и эукариот. Регуляторная и кодирующая субъединицы ...
Экология и биология даурской пищухи
Актуальность. Республика Тыва является составной частью Алтая-Саянского экорегиона, который входит в список, определенный Всемирным Фондом дикой природы ...
Что такое биология?
Биология (от греч. bios — жизнь и logos — учение), совокупность наук о живой природе. Предмет изучения — все проявления жизни: строение и функции ...
Системная биология - модели
системная биология - модели. потоковые линейное программирование эксперименты Виткуп Палссон – необычные источники Палссон – мутанты кинетические ...
Понятие популяции в экологии. основные свойства
Таким образом, народом в популяции выступают особи – совокупность особей. Особь – это отдельный организм (живое существо, индивид: лат.individuum ...
Новая биология
Концепция курса. ИУМК “Новая биология” предназначен для 6-9 классов средней общеобразовательной школы. Содержание курса отличается от традиционного ...
Важная биология в икт
Применение нашего компьютера:. моделирование биологических систем организация и хранение информации документооборот обучение экологические ГИС интернет-технологии. ...
Ботаника и биология
Г. Галилей (1564—1642). Развитие цитологии. Р. Гук (1635—1703). Антони ван Левенгук (1632—1723). Линзы Левенгука. Современные увеличительные приборы. ...
Наука биология
Наука биология. О чем? Когда? Для чего? Биология. Биология – это учение о жизни, или наука о живой природе. логос" - учение биос" - жизнь. Разделы ...
Наука биология и ее значение в нашей жизни
Что такое биология? Биология – наука о жизни, о живых организмах, обитающих на Земле. Что изучает биология? Биология изучает строение и жизнедеятельность ...

Общая биология
План:. 1.Общая биология- комплексная наука об общих закономерностях живой природы. 2.Общие признаки живого. 3.Уровни организации живой природы. 4.Роль ...
Веселая биология
Конкурсы. 1. Разминка «Биологические шарады» 2. Веселые загадки 3. Верните зверей в слова 4. Одни словом 5. Черный ящик 6. Ребусы. Биологические шарады. ...
Регенеративная биология и регенеративная медицина
РЕГЕНЕРАТИВНАЯ БИОЛОГИЯ как НАПРАВЛЕНИЕ ФУНДАМЕНТАЛЬНОЙ БИОЛОГИИ: РЕГЕНЕРАЦИЯ как БИОЛОГИЧЕСКИЙ ФЕНОМЕН -. ЖИВЫЕ ФОРМЫ ХАРАКТЕРИЗУЮТСЯ СВОЙСТВОМ – ...Советы как сделать хороший доклад презентации или проекта
- Постарайтесь вовлечь аудиторию в рассказ, настройте взаимодействие с аудиторией с помощью наводящих вопросов, игровой части, не бойтесь пошутить и искренне улыбнуться (где это уместно).
- Старайтесь объяснять слайд своими словами, добавлять дополнительные интересные факты, не нужно просто читать информацию со слайдов, ее аудитория может прочитать и сама.
- Не нужно перегружать слайды Вашего проекта текстовыми блоками, больше иллюстраций и минимум текста позволят лучше донести информацию и привлечь внимание. На слайде должна быть только ключевая информация, остальное лучше рассказать слушателям устно.
- Текст должен быть хорошо читаемым, иначе аудитория не сможет увидеть подаваемую информацию, будет сильно отвлекаться от рассказа, пытаясь хоть что-то разобрать, или вовсе утратит весь интерес. Для этого нужно правильно подобрать шрифт, учитывая, где и как будет происходить трансляция презентации, а также правильно подобрать сочетание фона и текста.
- Важно провести репетицию Вашего доклада, продумать, как Вы поздороваетесь с аудиторией, что скажете первым, как закончите презентацию. Все приходит с опытом.
- Правильно подберите наряд, т.к. одежда докладчика также играет большую роль в восприятии его выступления.
- Старайтесь говорить уверенно, плавно и связно.
- Старайтесь получить удовольствие от выступления, тогда Вы сможете быть более непринужденным и будете меньше волноваться.
Информация о презентации
Дата добавления:12 декабря 2018
Категория:Биология
Содержит:43 слайд(ов)
Поделись с друзьями:
Скачать презентацию


")






