Слайд 1Микробные объекты в биотехнологии
Принципы клеточной инженерии микроорганизмов
к.х.н., доцент кафедры микробиологии Герловский Денис Олегович
Минск, 2015

Слайд 2Искусственные ассоциации с микроорганизмами как способ модификации растительной клетки и растения В последние годы в области клеточной инженерии растительной клетки выделилось направление по созданию новых клеток и клеточных систем путем введения микроорганизмов в клетку или в популяции культивируемых клеток растений. Экспериментально создаваемые клеточные системы называют ассоциациями по аналогии с ассоциациями, формирующимися в природе между организмами разных видов. При этом исследования направлены на получение ассоциаций внутриклеточного (эндосимбиотического) или межклеточного (экзосимбиотического) типа. В первом случае проводят индуцированное введение микроорганизмов в изолированные протопласты высших растений. Во втором — совместно культивируют клетки или ткани растений с микроорганизмами. Хотя, как будет видно при дальнейшем изложении, исходно задаваемая в эксперименте локализация микроорганизмов — внутри клеток или в межклетниках тканей — не всегда сохраняется в процессе создания и культивирования таких систем.

Слайд 3При получении ассоциаций на основе изолированных протопластов или культивируемых клеток высших растений с микроорганизмами предполагается, что клетки или популяции клеток растений должны приобретать новые свойства, обусловленные присутствием в них клеток микроорганизмов. Возможность получения из изолированного протопласта клетки, если она сохранится у протопластов и после введения в них микроорганизмов, создает предпосылку для модификации клеток. Совместное культивирование растительных клеток и микроорганизмов позволило бы получать популяции растительных клеток с новыми свойствами, приобретенными в результате их взаимодействия с клетками микроорганизмов. И наконец, способность растительной клетки in vitro дать начало целому растению открывает возможность направленного изменения растений. Очевидно, последнее осуществимо при условии, что микроорганизмы, введенные внутрь клеток или в популяции клеток, сохранятся и в растениях, регенерированных из таких клеток.

Слайд 4Цели создания ассоциаций. Искусственные ассоциации клеток высших растений и микроорганизмов могут быть использованы для решения следующих фундаментальных научных проблем и практических задач. 1. Экспериментальная проверка теории эндосимбиотического происхождения эукариотной клетки, которое происходило, как полагают, через стадию экзосимбиоза. 2. Реконструирование отдельных стадий эволюционного процесса симбиогенеза. Возникновение ныне существующих симбиозов высших растений с микроорганизмами произошло в результате длительного и сложного эволюционного процесса. Воспроизведение природных симбиозов в лаборатории позволило бы понять механизмы возникновения и условия для формирования их заново в природе. 3. Моделирование и изучение на клеточном уровне природных симбиотических отношений (например, бобовых растений с клубеньковыми бактериями), имеющих большое практическое значение в обеспечении растений связанным азотом в природных экосистемах и в агрофитоценозах. 4. Повышение продуктивности растительной клетки при выращивании в культуре. 5. Улучшение экономически важных видов растений. Остановимся более подробно на возможности применения в биотехнологии искусственных ассоциаций с микроорганизмами, получаемых на основе культур растительных клеток или регенерированных из них растений.
 важных для медицины и ряда отраслей промышленности веществ. Одним из требований рентабельности производства на основе растительных клеток является возможность культивировани")
Слайд 5Повышение продуктивности культур растительных клеток. Растительные клетки в культуре проводят биосинтез (или биотрансформацию) важных для медицины и ряда отраслей промышленности веществ. Одним из требований рентабельности производства на основе растительных клеток является возможность культивирования на простых по составу питательных средах. Между тем культуры растительных клеток требуют присутствия в среде витаминов, фитогормонов, аминокислот, сахарозы и других веществ. Серьезным ограничением практического использования культур растительных клеток является энергетическое обеспечение процессов биосинтеза. Растительные клетки в культурах гетеротрофны или обладают ограниченной способностью к фотосинтезу. Представляется маловероятным совместить в культивируемой клетке способность к собственному фотосинтезу и биосинтезу видоспецифических продуктов. Одним из решений данной проблемы могло бы быть введение в эти культуры микроорганизмов, синтезирующих субстраты для роста растительных клеток или предшественники для биосинтеза.

Слайд 6Особое значение при этом, очевидно, имеет введение в гетеротрофно растущие клетки или культуры клеток растений фототрофных микроорганизмов. Последние могли бы обеспечить растительные клетки продуктами фотосинтетической фиксации углерода, что позволило бы упростить состав среды. В литературе отмечается недостаточное использование в биотехнологии принципов организации природных биологических систем, в частности, основанных на взаимоотношениях организмов разных видов. В микробиологии уже накоплен положительный опыт смешанного культивирования. В результате изучения смешанных культур микроорганизмов (выделенных из природы или созданных искусственно) выяснилось, что можно проводить более эффективно (по сравнению с монокультурами) накопление биомассы, кооперативный биосинтез конечных продуктов или трансформацию в нужном направлении исходного субстрата. Такие системы находят все большее применение в микробиологической промышленности и могут быть использованы для очистки сточных вод, биосинтеза белка (ферментов) и биологически активных веществ, таких, как ауксины, витамины, антибиотики. Считается, что в биотехнологии найдут применение смешанные популяции, разнообразные по своему составу, начиная от комбинаций нескольких штаммов одного вида микроорганизма и кончая сочетаниями представителей разных царств — животного и растительного.

Слайд 7Улучшение сельскохозяйственных растений. Другая возможность использования в биотехнологии ассоциаций клеток высших растений с микроорганизмами состоит в получении растений, способных к фиксации молекулярного азота. Повышение урожайности сельскохозяйственных растений в значительной степени зависит от обеспечения их связанным азотом. Растения получают его двумя путями. Один из них — потребление внесенных в почву химических азотных удобрений, на получение которых затрачивается огромная энергия. Растения же используют азотные удобрения частично — от 30 до 50% вносимого количества. Продукты разложения азотных удобрений в почве образуют токсичные соединения, загрязняющие окружающую среду. Другим источником является биологическая фиксация молекулярного азота, которую осуществляют многие группы микроорганизмов. Она безвредна для человека и окружающей среды (в том объеме, в котором она осуществляется в агрофитоценозах и в естественных экосистемах). Большая часть азота в природе фиксируется симбиотическими азотфиксаторами. Они используют продукты фотосинтеза макросимбионта для покрытия энергетических затрат на фиксацию азота и передают связанный азот растению. Способность формировать азотфиксирующие симбиозы, однако, приобрели в процессе эволюции только определенные виды растений и микроорганизмов. Все это обусловило интерес исследователей к разработке проблемы повышения доли биологического азота в питании растений.

Слайд 8Практика сельскохозяйственного производства поставила вопрос придания способности к азотфиксации экономически важным видам растений, которым это не свойственно. В решении этой задачи предлагаются разные подходы. Растения искусственно инокулируют азотфиксирующими бактериями путем введения микроорганизмов в почву и ризосферу. Однако получаемый при этом эффект повышения урожая растений во многих случаях связан не с фиксацией бактериями азота, а с увеличением под влиянием бактерий числа корней и в результате более активным поглощением растениями ионов и воды. Кроме того, отмечается низкая выживаемость и вытеснение интродуцируемых чистых культур микроорганизмов естественной почвенной микрофлорой. Поэтому этот подход представляется бесперспективным.
 от микроорганизмов, фиксирующих азот, непосредственно в злаковые или другие экономически важные виды растений. По мнению специалистов, эта п")
Слайд 9Другой способ — применение методов генетической инженерии для создания растений, способных к азотфиксации. Предлагается переносить гены азотфиксации (nif-гены) от микроорганизмов, фиксирующих азот, непосредственно в злаковые или другие экономически важные виды растений. По мнению специалистов, эта процедура методически вполне осуществима; nif-ген может быть введен в протопласты растений с помощью определенных векторов (таких, как плазмиды бактерий, патогенных для растений, или вирусы растений). Последующее культивирование протопластов и их регенерация до целых растений позволили бы получить особи, которые несли бы введенную генетическую информацию в своих клетках и, возможно, в последующих поколениях благодаря передаче через семена. Однако, несмотря на принципиальную возможность применения такого подхода, привлечение методов генетической инженерии вряд ли позволит в ближайшее время перейти к задаче создания азотфиксирующих растений. Основные сложности состоят в следующем: требуется разработка методов введения nif-генов в растительную клетку, их репликации и экспрессии там; у высших растений отсутствуют системы, которые осуществляли бы энергообеспечение фермента азотфиксации — нитрогеназы (процесс азотфиксации связан с затратой большого количества клеточной энергии); растительная клетка не обладает соответствующими системами транспорта и запасания в высокой концентрации ионов железа и молибдена, необходимых для синтеза нитрогеназы; наконец, она не имеет системы защиты нитрогеназы от инактивации кислородом. Последнее обстоятельство считают главным лимитирующим фактором в экспрессии nif-генов при введении их в аэробные организмы.

Слайд 10Ограничения указанных двух подходов по приданию растениям способности к фиксации молекулярного азота могут быть хотя бы частично преодолены при введении целых клеток азотфиксирующих микроорганизмов в растение. Речь идет об альтернативном способе решения обсуждаемой проблемы, связанном с конструированием новых искусственных ассоциаций. Такие системы должны быть основаны на симбиотических взаимоотношениях растительного и микробного партнеров. В этих экспериментах должны учитываться особенности организации природных азотфиксирующих симбиозов, а именно: 1) сохранение целостности обоих партнеров; 2) интеграция партнеров в пределах организма макросимбионта; 3) относительная обособленность микросимбионта, компартментация его в специальных структурах (например, локализация клубеньковых бактерий в клубеньках у бобовых растений, цианобактерии Anabaena azollae — в полости листа папоротника Azolla). Азотфиксация в таких искусственно создаваемых системах осуществлялась бы за счет функционирования азотфиксирующих микроорганизмов в клетках или тканях растений.

Слайд 11Достижение этой цели возможно с использованием техники клеточной инженерии. Введение азотфиксирующих бактерий в клетки или популяции культивируемых клеток небобовых растений позволяет испытать большое число сочетаний партнеров и выбрать устойчивые ассоциации на уровне культивируемых клеток. В процессе культивирования возможна адаптация партнеров к совместному существованию, аналогичная тем изменениям, которые приобретают компоненты природных симбиотических ассоциаций по сравнению со свободноживущими формами. Последующее получение растения в результате индукции органогенеза при условии сохранения клеточных взаимодействий, складывающихся в клеточных системах, позволило бы решать поставленную задачу — инкорпорирования бактериального симбионта в ткани (клетки) растения, интеграции их в клеточные ансамбли хозяина при сохранении интактности вводимого симбионта. Очевидны преимущества введения в клетки и ткани растений целых клеток азотфиксаторов. Защита нитрогеназы от кислорода, выделяемого растением и необходимого для его роста, в таком случае будет осуществляться с помощью механизмов, которыми обладают бактериальные клетки. Особого внимания заслуживает использование в экспериментах азотфиксаторов, способных к фотосинтезу. Такие организмы могли бы обеспечить нитрогеназу энергией, получаемой в процессе собственного фотосинтеза. В случае применения гетеротрофных азотфиксаторов остро стоит вопрос о том, сможет ли растение в ассоциации с бактериями образовать достаточное количество продуктов фотосинтеза, чтобы обеспечить энергетические потребности нитрогеназы симбионта без ущерба для своего роста и ухудшения потребительских качеств растения (по содержанию углеводов, белка и т. д.).

Слайд 12Введение микроорганизмов в изолированные протопласты растений Партнеры создаваемых ассоциаций. С целью получения ассоциаций внутриклеточного типа предпринимались неоднократные попытки вводить микроорганизмы в изолированные протопласты разных видов высших растений, среди которых представлены злаковые и другие сельскохозяйственные культуры, не имеющие естественных симбионтов. Протопласты выделяют из тканей растений или клеток суспензионных культур ферментативным способом. В изолированные протопласты растений вводили микроорганизмы разных систематических групп: бактерии, дрожжи, цианобактерии (прежнее их название — синезеленые водоросли), органеллы (называемые цианеллами) водоросли Glaucocystis nostochinearum и криптомонадоподобного простейшего Cyanophora paradoxa. Цианеллы представляют собой эндосимбиотические цианобактерии с отсутствующей или редуцированной клеточной стенкой. Используемые в экспериментах микроорганизмы отличались своими метаболическими возможностями. Зеленые водоросли, все цианобактерии и цианеллы осуществляют фотосинтез;

Слайд 13бактерии R. leguminosarum, A. brasilense, цианобактерии Gloeocapsa, A. variabilis и N. macrozamia — азотфиксаторы. Цианобактерия A. variabilis — факультативный фототроф и способна к гетеротрофному росту в темноте. Наряду с диким штаммом в протопласты вводили также ауксотрофные мутанты A.variabilis, испытывающие потребность в определенных факторах роста, которые они могли бы получать от растительной клетки. Такой прием применяют с целью установления возможной зависимости эндосимбионта от роста и метаболизма клетки хозяина.
Слайд 14

Слайд 15Механизмы проникновения микроорганизмов в протопласты. Введение микроорганизмов в изолированные протопласты растений проводят при воздействии на них специальными индуцирующими факторами. Микроорганизмы могут поглощаться протопластами или сливаться с ними. Протопласты гороха поглощали клетки Rhizobium непосредственно в процессе ферментативного разрушения клеточных стенок клеток мезофилла листа при получении изолированных протопластов. Во всех остальных случаях в качестве индуктора поглощения или слияния применяли полиэтилеигликоль (ПЭГ), который известен как агент, повышающий частоту слияния протопластов высших растений. Поглощение растительными протопластами микроорганизмов (кроме зеленых водорослей) может происходить путем эндо цитоза, т.е. с помощью инвагинации плазмалеммы. При действии ПЭГ микроорганизмы прикрепляются к плазмалемме, образуется инвагинация плазмалеммы вокруг микроорганизма с последующим замыканием конца инвагинации путем слияния плазмалеммы. В результате образования таким образом везикула с микроорганизмом высвобождается в цитоплазму протопласта.
 с протопластами моркови индуцируется ПЭГ по аналогии со слиянием протопластов высших растений. В этом случае происходит интеграция цитоплазматических мембран растительных протопластов и водорослей, а органеллы водорослей высвоб")
Слайд 16Слияние зеленых водорослей (протопластов и мутанта без клеточной стенки) с протопластами моркови индуцируется ПЭГ по аналогии со слиянием протопластов высших растений. В этом случае происходит интеграция цитоплазматических мембран растительных протопластов и водорослей, а органеллы водорослей высвобождаются в цитоплазму протопластов, но не окружаются плазмалеммой протопласта. В связи с проблемой сохранения интактности вводимых в протопласты органелл или микроорганизмов тот и другой способы имеют свои недостатки. Поглощение путем эндоцитоза приводит к изолированию вводимых в протопласт объектов в везикулах из плазмалеммы протопласта. Поскольку эти везикулы могут сливаться с лизосомальным аппаратом растительного протопласта, существует опасность, что это приведет к разрушению вводимого чужеродного материала. При слиянии происходит интеграция мембраны протопласта растения и микроорганизма, нарушение целостности микроорганизма и освобождение его органелл внутрь растительного протопласта. Обнаруженное для зеленых водорослей слияние с протопластами может рассматриваться, таким образом, как способ введения в растительную клетку не целых микроорганизмов, а интактных органелл. В качестве альтернативного пути, позволяющего преодолеть недостатки обоих рассматриваемых способов, предлагается заключать микроорганизмы в искусственные мембраны — липосомы. Этот прием уже был использован в опытах по введению в протопласты лука одноклеточных цианобактерий, заключенных в липидные капли. Преимущества данного методического приема видятся в том, что искусственные мембраны будут сливаться с плазмалеммой протопласта, освобождая таким образом интактные клетки микроорганизмов в цитоплазму протопласта.

Слайд 17
Слайд 18Помимо использования ПЭГ в качестве индуктора поглощения протопластами микроорганизмов, можно отметить успешное применение в этих целях поливинилового спирта (ПВС), который также известен как индуктор слияния протопластов. При воздействии ПВС протопласты Vinca rosea поглощали сферопласты Agrobacterium tumefaciens (S. Hasezama et al., 1983), клетки и сферопласты Escherichia coli (С. Matsui et al., 1983) в экспериментах, проводимых с целью генетической трансформации протопластов. В этих опытах ПВС не оказывал токсического эффекта на протопласты, который был отмечен в сравнительных опытах с использованием ПЭГ.

Слайд 19Характеристика продуктов поглощения и слияния. Системы, полученные путем поглощения протопластами микроорганизмов или слияния с ними, обладали ограниченной способностью к выживанию или культивированию. В протопластах кукурузы обнаружено быстрое движение цитоплазмы через 14 ч после поглощения ими клеток цианобактерии, что служит признаком их жизнеспособности. В некоторых случаях протопласты, содержащие клетки или органеллы микроорганизмов, в процессе культивирования регенерировали клеточные стенки, и только в случае поглощения органелл С. reinhardii были способны к последующему делению. Клетки табака с внутриклеточной A. variabilis никогда не делились и через 5 сут культивирования разрушались. Есть отдельные данные о сохранении интактности клеток и органелл микроорганизмов внутри протопластов. В протопластах гороха в везикулах наблюдали делящиеся клетки Rhizobium, однако осталось неясным, начали они деление до или после поглощения их протопластом. Морфология бактерий свидетельствует об их интактном состоянии. Не происходило деления цианобактерии A. variabilis в протопластах табака в процессе их культивирования, хотя цианобактерии сохраняли видимую интактность. Ни один из акусотрофных мутантов этой цианобактерии не имел преимуществ в формировании внутриклеточных систем, гибель которых происходила с той же скоростью, что и систем, образуемых с диким штаммом. Органеллы зеленых водорослей в растительных протопластах в процессе культивирования деградировали, и только хлоропласты обнаруживались в цитоплазме клеток моркови через 10 сут.
 получали из бо")
Слайд 20Введение микроорганизмов в популяции культивируемых клеток растений. Партнеры ассоциаций и принципы их создания. При получении ассоциаций на основе культивируемых клеток и тканей используется разнообразный видовой набор растительных объектов и микроорганизмов. Культуры клеток (тканей) получали из бобовых, злаковых, овощных и других экономически важных растений, в том числе — продуцентов природных веществ. Микроорганизмы представлены в основном азотфиксирующими формами, среди которых симбиотические (клубеньковые) и свободноживущие (Azotobacter, Azospirillum) бактерии, а также несимбиотические цианобактерии. Кроме того, для получения ассоциаций с каллусными культурами использовали зеленую водоросль Chlorella и разные виды грибов. Цели создания ассоциаций с отдельными микроорганизмами существенно различаются так же, как и критерии, на основе которых оценивается возникновение ассоциативных взаимодействий в системах. Общим принципом для создания ассоциаций является совместное культивирование клеток и каллусных тканей растений с микроорганизмами. При этом способы введения микроорганизмов в систему для совместного выращивания могут быть разными: внесение клеток микроорганизмов непосредственно в каллусную или суспензионную культуру растения (смешанные культуры); высев на поверхность агаризованной среды рядом с каллусом таким образом, чтобы рост микроорганизмов происходил без соприкосновения (совместные культуры); разделение бактериальных клеток и клеток суспензионной или каллусной культуры специальными фильтрами (мембранами), обеспечивающими обмен продуктами метаболизма, но не допускающими контактов между клетками партнеров.

Слайд 21

Слайд 22Ассоциации с клубеньковыми бактериями. Основная задача при получении ассоциаций с клубеньковыми бактериями первоначально состояла в создании упрощенных модельных систем для изучения симбиотических взаимоотношений бобового растения с клубеньковыми бактериями. Впервые каллусная ткань сои была инфицирована клетками Rhizobium, вызывающими лигнификацию и дифференцировку растительных клеток. Позже была получена ассоциация, в которой клубеньковые бактерии формировали подобие инфекционных нитеи и бактероиды в каллусных клетках сои. Цитологическое изучение показало сходство процессов инфицирования в искусственной ассоциации и при формировании корневого клубенька в целом растении. Бактерии инфицировали до 10% клеток каллуса, обнаруживаясь в цитоплазме в везикулах, образованных, очевидно, плазмалеммой растительной клетки. В клетке они размножались, заполняя цитоплазму и образуя бактероиды, как и в клубеньке растения. В ассоциации с каллусными клетками клубеньковые бактерии проявляли нитрогеназную активность (НГА), которая отсутствует у них в чистой культуре. Это свидетельствует об азотфиксации клубеньковыми бактериями в ассоциациях с каллусными клетками, которая составляла 1 % от активности азотфиксации в клубеньках растения. В последующих исследованиях была подтверждена индукция НГА у клубеньковых бактерий культурами клеток (тканей) бобовых растений. Существенными для понимания взаимодействия макро- и микроорганизма в природных симбиозах бобовых оказались сообщения об индукции НГА у Rhizobium тканевыми культурами небобовых растений —пшеницы, рапса, табака и других видов.

Слайд 23Таким образом, получение азотфикеирующих ассоциаций при взаимодействии клубеньковых бактерий с клетками небобовых растений приводит к выводу о принципиальной возможности распространения симбиоза с клубеньковыми бактериями на небобовые культуры. На этом основании различные комбинации эукариотных клеток и азотфиксирующих микроорганизмов могут быть использованы в экспериментах in vitro для выяснения высокой специфичности природных симбиозов и понимания причин, по которым симбиоз в природе избирательно возник только у некоторых групп растений.
 бобовых и небобовых растений с симбиотическими (клубеньковыми) бактериями послужили основой для получения ассоциаций с несимбиотическими азотфиксаторами — бактериями родов Azotobact")
Слайд 24Ассоциации с азотфиксирующими свободноживущими бактериями. Эксперименты по созданию ассоциаций культур клеток (тканей) бобовых и небобовых растений с симбиотическими (клубеньковыми) бактериями послужили основой для получения ассоциаций с несимбиотическими азотфиксаторами — бактериями родов Azotobacter и Azospirillum. При создании искусственных ассоциаций на основе культур растительных тканей ставилась задача получить эффективные азотфиксирующие системы, растущие на среде без связанного азота, с перспективой регенерации растений. В этих целях использовали сочетания культур растительных тканей с бактериями A. vinelandii и A. brasilense. Была обнаружена видовая специфичность растительных клеток в формировании ассоциаций с этими бактериями. Каллусные культуры табака, проса и Eremochloa ophiuroides не выживали при инокуляции их Azospirillum. Каллусные культуры быстро «обрастали» бактериями, погибая через 4 недели после инокуляции. Напротив, при инокуляции той же бактерией ткани сахарного тростника была получена ассоциация, которую удалось субкультивировать на протяжении 18 мес. Стабильная ассоциация формировалась только на среде с низким содержанием связанного азота или полностью лишенной его. Бактерии проявляли НГА независимо от того, формировали они ассоциации с растительными тканями или последние погибали.

Слайд 25Как Azotobacter, так и Azospirillum в соответствующих ассоциациях были локализованы на поверхности или в межклетниках каллусной ткани и никогда не проникали в живые растительные клетки. Попытка регенерации растений из ассоциации каллуса моркови с Azotobacter была неудачной. В ассоциации ткани сахарного тростника и Azospirillum при инкубации на свету были регенерированы зеленые побеги, которые не отличались от побегов, полученных в контрольной каллусной культуре. Они были способны к формированию корней и росту в почве. Однако ни в побегах, ни в корнях бактериальные клетки не обнаруживались. Не- возможность проникновения бактерий в побеги в процессе органогенеза в смешанной культуре связывают с анатомическими особенностями каллуса сахарного тростника. Данная каллусная культура имеет очень уплотненные области, характеризующиеся отсутствием межклетников. Рост бактерий происходит в поверхностных участках каллусных агрегатов, что сопровождается отмиранием растительных клеток. Благодаря отсутствию межклетников бактерии не проникают в глубь этих агрегатов, где сохраняется жизнеспособная растительная ткань. Образование меристемоидов происходит в уплотненных участках каллуса, лишенных межклетников, что препятствует включению бактерий в формирующиеся побеги.

Слайд 26Ассоциация с зеленой водорослью. Инокуляция каллуса моркови одним из штаммов Chlorella и культивирование системы на дефицитной по азоту среде на свету приводила к выживанию каллуса в течение нескольких месяцев, в то время как контрольный каллус погибал. Клетки водоросли росли на поверхности каллуса и проникали в межклетники. В инокулированном каллусе обнаружен высокий процент интактных клеток, и большинство из них, очевидно, были живыми. Клетки другого штамма водоросли не росли совместно с каллусом, и в их присутствии обнаружена очень низкая доля интактных клеток моркови. На основании проведенных экспериментов сделан вывод об обеспечении клеток моркови азотистыми веществами, поступающими от водоросли, при культивировании системы на среде с дефицитом азота. Ассоциации с грибами. Способ совместного культивирования ткани руты с различными грибами был применен в качестве нового, нетрадиционного подхода к повышению биосинтеза видоспецифических продуктов, образуемых растительными клетками in vitro (В. Wolters, U. Eilert, 1982). Совместную культуру получали таким образом, чтобы каллус и мицелий гриба не соприкасались, т. е. на биосинтез влияли диффундирующие через агар выделения гриба.

Слайд 27Различные виды грибов по-разному воздействовали на рост каллуса руты и биосинтез клетками алкалоидов акридонового типа. При этом грибы вызывали подавление или усиление роста каллуса по сравнению с контролем, сопровождающееся повышением видоспецифических биосинтезов. Стимулирующее действие на биосинтез оказывала также культуральная жидкость грибов. Можно выделить 3 группы фактов, полученных в результате изложенных здесь экспериментов, которые важны для развития работ по конструированию новых клеточных систем. 1. Показана возможность получения смешанных клеточных систем, растущих на среде без связанного азота, по-видимому, за счет бактериальной фиксации молекулярного азота. 2. Выявлен нормальный ход органогенеза в растительной ткани в присутствии бактерий, хотя, к сожалению, в данных опытах бактерии и не включались в растения. 3. Установлено значительное повышение видоспецифических биосинтезов растительными клетками под влиянием микроорганизмов. Разработанные подходы могут быть, таким образом, использованы для получения систем, моделирующих природные симбиотические отношения, а также для дальнейшего экспериментирования в целях улучшения свойств растительной клетки in vitro или целого растения.

Слайд 28Особенности цианобактерий в качестве партнера растительных клеток в искусственных ассоциациях. В исследованиях по получению ассоциаций на основе изолированных протопластов были сделаны попытки введения цианобактерий в изолированные протопласты. Кроме того, выполнены единичные работы по совместному культивированию цианобактерий с растительными тканями. При этом было показано сохранение интактности клеток каллуса моркови в условиях освещенности на среде, дефицитной по азоту, при совместном культивировании с азотфиксирующими цианобактериями Anabаеnа и Plestonema. В смешанной культуре с каллусом табака выращивали так же азотфикснруюшие цианобактерии. На среде без азота в отсутствие цианобактерий каллус табака погибал. Проведена селекция штаммов цианобактерий, способных к совместному росту с каллусом в течение 20 мес, у цианобактерий зарегистрирована НГА.
Слайд 29Спасибо за внимание
 Слайд 1
Слайд 1 Слайд 2
Слайд 2 Слайд 3
Слайд 3 Слайд 4
Слайд 4 Слайд 5
Слайд 5 Слайд 6
Слайд 6 Слайд 7
Слайд 7 Слайд 8
Слайд 8 Слайд 9
Слайд 9 Слайд 10
Слайд 10 Слайд 11
Слайд 11 Слайд 12
Слайд 12 Слайд 13
Слайд 13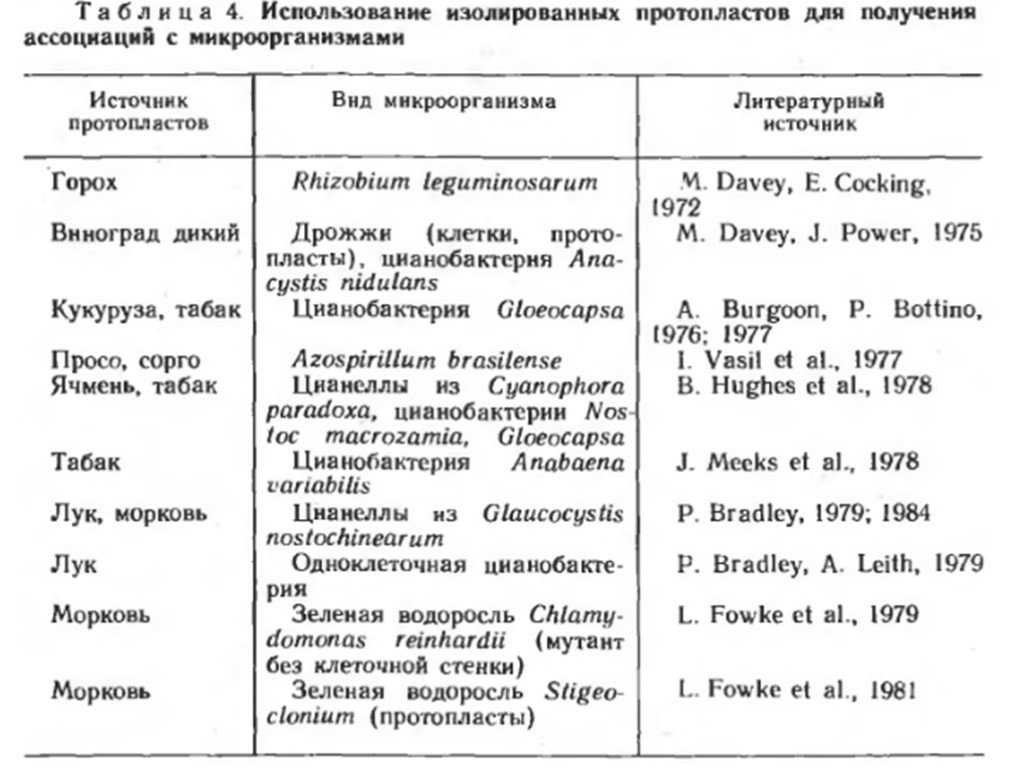 Слайд 14
Слайд 14 Слайд 15
Слайд 15 Слайд 16
Слайд 16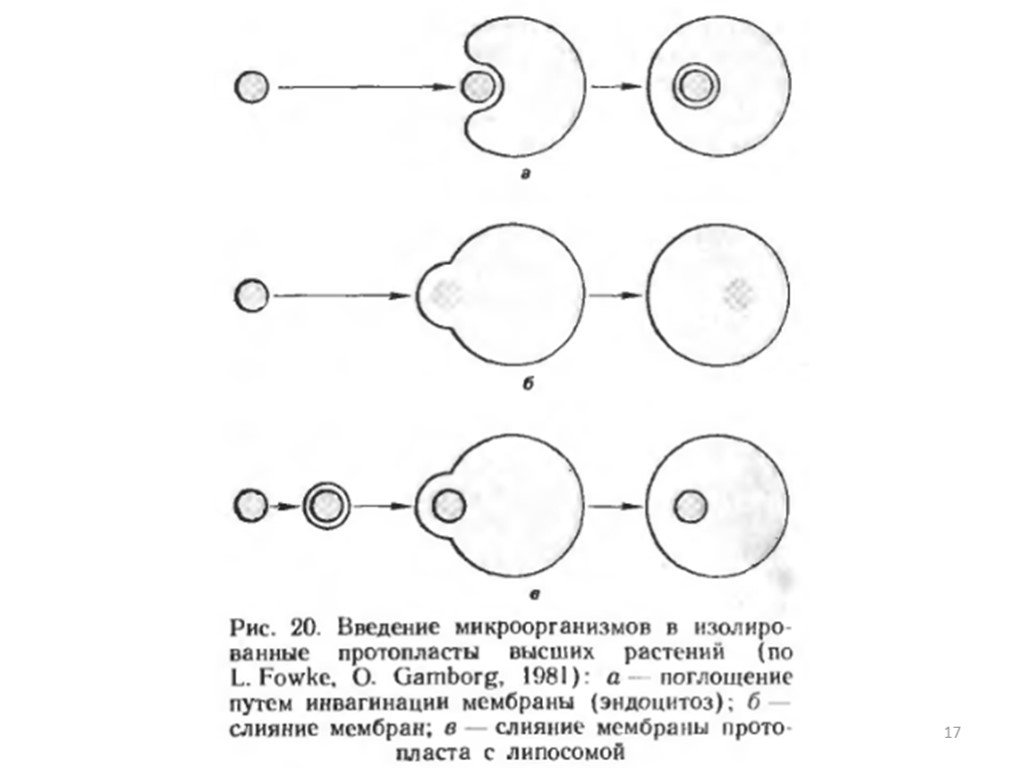 Слайд 17
Слайд 17 Слайд 18
Слайд 18 Слайд 19
Слайд 19 Слайд 20
Слайд 20 Слайд 21
Слайд 21 Слайд 22
Слайд 22 Слайд 23
Слайд 23 Слайд 24
Слайд 24 Слайд 25
Слайд 25 Слайд 26
Слайд 26 Слайд 27
Слайд 27 Слайд 28
Слайд 28 Слайд 29
Слайд 29













