Слайд 1Ведение в обмен веществ. Хромопротеиды
Кафедра биологической химии (биохимии) 20 сентября 2013
СТОМАТОЛОГИЧЕСКИЙ ФАКУЛЬТЕТ (ОЧНО-ЗАОЧНОЕ ОТДЕЛЕНИЕ) НА ТРЕТИЙ СЕМЕСТР 2013/2014уч.г.
Слайд 2СОДЕРЖАНИЕ ЛЕКЦИИ
- Понятие о катаболизме и анаболизме. - Основы питания. - Незаменимые компоненты пищи: *аминокислоты *минеральные вещества *витамины *витаминоподобные факторы - Хромопротеиды. - Гемоглобин, синтез гемоглобина. - Распад гемоглобина, образование пигментов желчи, кала и мочи. - Нарушение пигментного обмена.
Слайд 3Понятие о катаболизме и анаболизме.
Метаболизм (metabole — греч. измене- ние, превращение) — это совокупность процессов превращения веществ и энергии в организме, происходящих с участием ферментов. В наиболее употребительном значении термин «метаболизм» равнозначен «обмену веществ». В точном смысле «метаболизм» означает промежуточный обмен, т. е. превращение веществ внутри клеток с момента их поступления до образования конечных продуктов. Вещества, участвующие в метаболизме, называются метаболитами

Слайд 4Функции метаболизма
1. Обеспечение организма энергией, полученной при расщеплении богатых энергией пищевых веществ или путем преобразования энергии солнца. 2. Превращение пищевых молекул в предшественники, которые используются в клетке для биосинтеза собственных макромолекул. 3. Сборка макромолекулярных (биополимеры) и надмолекулярных структур живого организма, т. е. пластическое и энергетическое поддержание его структуры. 4. Синтез и разрушение биомолекул, выполняющих специфические функции в организме (мембранные липиды, внутриклеточные посредники и пигменты).
Слайд 5Фазы метаболизма
Метаболизм складывается из двух фаз — катаболизма и анаболизма. Катаболизм — это ферментативное расщепление крупных пищевых или депонированных молекул (углеводов, липидов, белков) до более простых (лактат, Н2О, СО2, NH3) с выделением энергии и запасанием ее в виде АТФ или восстановительных эквивалентов (НАДН, НАДФН, ФАДН2).
, Первая стадияпревращения пищевых молекул протекает в желудочно-кишечном тракт")
Слайд 6Катаболизм включает 3 стадии. 1 стадия — превращение полимеров в мономеры (крахмал и гликоген — в глюкозу, белки — в аминокислоты, триацилглицеролы — в жирные кислоты и глицерол, нуклеиновые кислоты — в нуклеотиды и т.д.), Первая стадияпревращения пищевых молекул протекает в желудочно-кишечном тракте и называется перевариванием. 2 стадия (специфические пути катаболизма) — мономеры превращаются в общие промежуточные продукты — пируват и ацетил-КоА. 3 стадия (общий путь катаболизма) — окисление ацетильной группы ацетил-КоА до СО2 и Н2О. 3 стадия катаболизма включает: а) цикл трикарбоновых кислот б) цепи переноса электронов в) окислительное фосфорилирование.

Слайд 7Анаболизм — ферментативный синтез крупных полимерных молекул из простых предшественников с затратой АТФ или восстановительных эквивалентов НАДН, НАДФН и ФАДН2. Стадии анаболизма: 1стадия — третья стадия катаболизма, т. е. цикл трикарбоновых кислот; 2 стадия — образование мономеров по реакциям, обратным реакциям катаболизма; 3 стадия — синтез полимеров из мономеров. Амфиболические пути расположены в точках переключения метаболизма и связывают анаболизм и катаболизм.
Слайд 8
Слайд 9Амфиболическим путем метаболизма является цикл трикарбоновых кислот
Слайд 10Анаболизм и катаболизм не являются простым обращением реакций. Катаболические и анаболические пути должны отличаться хотя бы одной из ферментативных реакций, чтобы регулироваться независимо. Углеводы, жирные кислоты и большинство аминокислот окисляются в конечном счёте через цикл лимонной кислоты до СО2 и Н2О. Прежде, чем эти вещества вовлекаются в заключительный этап катаболизма, их углеродный скелет превращается в двухуглеродный фрагмент в форме ацетил-КоА. Именно в этой форме большая часть «топливных» молекул включается в цикл лимонной кислоты. Ацетил-КоА образуется в специфических реакциях катаболизма жирных кислот и некоторых аминокислот Однако главным источником ацетил-КоА служит пировиноградная кислота, образующаяся в реакциях катаболизма глюкозы и некоторых аминокислот
 включает 11 реакций; обратный процесс — синтез глюкозы из лактата (глюконеогенез) включает 8 обратимых реакций и 3 дополнительные реакции с новыми наборами ферментов. Именно на этих стадиях за счет направленного изменения")
Слайд 11Например, специфический путь распада глюкозы до лактата (анаэробный гликолиз) включает 11 реакций; обратный процесс — синтез глюкозы из лактата (глюконеогенез) включает 8 обратимых реакций и 3 дополнительные реакции с новыми наборами ферментов. Именно на этих стадиях за счет направленного изменения активности ферментов регулируются суммарные скорости распада и синтеза глюкозы.

Слайд 12Несмотря на то, что обе стороны метаболизма сопряжены между собой во времени и пространстве, они строго локализованы в отдельных органеллах клетки и образуют самостоятельные метаболические пути. Локализация основных путей метаболизма в органеллах клетки отражена в следующей таблице:
Если бы эти пути совпадали или отличались лишь направлением процесса, то в обмене возникали бы бесполезные, так называемые фитильные циклы. Образование таких циклов является причиной ряда заболеваний, при которых происходит бесполезный круговорот метаболитов вследствие реакций, протекающих в патологическом режиме. Важнейшая черта биохимической формы движения материи заключается в саморегуляции совокупности биохимических превращений, образующих механизм, который обладает свойством обратной связи (положительной или отрицательной). Необходимо отметить, что каждая из биохимических реакций данного механизма в отдельности не обладает способностью к саморегулированию.

Слайд 13Основы питания.
Сбалансированный пищевой рацион человека должен включать множество различных компонентов. К ним относятся белки, углеводы, жиры, витамины и минеральные вещества, а также вода. В зависимости от режима питания относительное и абсолютное содержание питательных веществ в рационе может сильно варьировать. Поскольку некоторые питательные вещества являются незаменимыми (эссенциальными — жизненно важными), они должны поступать в организм регулярно. Минимальная суточная потребность в различных питательных веществах дана в рекомендациях ВОЗ (Всемирная организация здравоохранения) и национальных организаций по охране здоровья.
Слайд 14Основы питания
Энергетические потребности Энергетические потребности организма человека зависят от возраста, пола, массы, состояния здоровья и физической активности. При этом рекомендуется, чтобы примерно половина суточного энергообеспечения поступала в виде углеводов, не более трети с жирами, а остальное с белками.

Слайд 15Минимальная суточная потребность в белке составляет у мужчин 37 г, у женщин 29 г, однако рекомендованные нормы потребления почти вдвое выше. Еще выше нормы потребления для женщин в период беременности и кормления ребенка. При оценке пищевых продуктов важно также учитывать качество белка. При отсутствии или низком содержании незаменимых аминокислот белок считается малоценным. Соответственно такие белки должны потребляться в большем количестве. Так, белки бобовых содержат мало метионина, а белки пшеницы и кукурузы характеризуются низким содержанием лизина. Напротив, животные белки (исключая коллагены и желатину) относятся к полноценным пищевым продуктам. Белки являются жизненно необходимыми компонентами питания, поскольку они служат источником незаменимых аминокислот, которые не могут синтезироваться в организме человека. Некоторые аминокислоты, в том числе цистеин и гистидин, хотя и не относятся к незаменимым, необходимы для нормального роста и развития. Многие аминокислоты в пищевых продуктах взаимозаменяемы. Так, незаменимая аминокислота тирозин может образовываться в организме человека путем гидроксилирования фенилаланина, а цистеин может получаться из метионина.
, дисахаридов (в молоке и всех сладостях, содержащих сахарозу), а также полисахаридов растительного (крахмал) и животного (гликоген) происхождени")
Слайд 16Углеводы служат общим и легко утилизируемым источником энергии. В пищевых продуктах углеводы присутствуют в виде монасахаридов (например, в меде и фруктах), дисахаридов (в молоке и всех сладостях, содержащих сахарозу), а также полисахаридов растительного (крахмал) и животного (гликоген) происхождения. Несмотря на то, что углеводы вносят существенный вклад в энергообеспечение организма, они не считаются незаменимыми питательными веществами. Жиры — наиболее важный источник энергии. По энергетической ценности они вдвое превосходят белки и углеводы. Кроме того, жиры выполняют функции переносчиков жирорастворимых витаминов, а также служат источником полиненасыщенных жирных кислот, необходимых для биосинтеза эйкозаноидов.

Слайд 17Минеральные вещества В количественном отношении наиболее важным неорганическим компонентом пищи является вода. У взрослого человека суточная потребность в воде составляет примерно 2,4 л. Эта цифра включает воду, поступающую в организм с твердой и жидкой пищей, в виде напитков, а также воду, образующуюся в дыхательной цепи. Жизненно необходимые элементы подразделяются на макроэлементы (суточная потребность >100 мг) и микроэлементы (суточная потребность
. Среднесуточная потребность взрослого человека в указанных элементах приведена в колонке 4. У детей и женщин в период беременности и кормления ребенка, а также у больных потребность в")
Слайд 18В таблице приведено среднее содержание минеральных веществ в организме взрослого человека (в расчете на массу 65 кг). Среднесуточная потребность взрослого человека в указанных элементах приведена в колонке 4. У детей и женщин в период беременности и кормления ребенка, а также у больных потребность в микроэлементах обычно выше. Так как многие элементы и вода могут запасаться в организме, отклонение от суточной нормы компенсируется во времени. Вода запасается во всех тканях организма, кальций — в форме апатита костной ткани (см. с. 332), иод — в составе тиреоглобулина в щитовидной железе, железо — в составе ферритина и гемосидерина в костном мозге, селезенке и печени. Местом хранения многих микроэлементов служит печень. Обмен минеральных веществ контролируется гормонами.
Слайд 19

Слайд 20Количество минеральных веществ, абсорбированных из пищи, как правило, зависит от метаболических потребностей организма и в ряде случаев от состава пищевых продуктов. В качестве примера влияния состава пищи можно рассмотреть кальций. Всасыванию ионов Ca2+ способствуют молочная и лимонная кислоты, в то время как фосфат-ион, оксалат-ион и фитиновая кислота ингибируют всасывание кальция из-за комплексообразования и образования плохо растворимых солей (фитин). Дефицит минеральных веществ — явление не столь редкое: оно возникает по различным причинам, например из-за однообразного питания, нарушения усвояемости, при различных заболеваниях. Недостаток кальция может наступить в период беременности, а также при рахите или остеопорозе. Хлородефицит наступает из-за большой потери ионов Сl- при сильной рвоте. Из-за недостаточного содержания иода в пищевых продуктах во многих районах Центральной Европы распространенным явлением стали иододефицитные состояния и зобная болезнь. Дефицит магния может возникать из-за диареи или из-за однообразного питания при алкоголизме. Недостаток в организме микроэлементов часто проявляется нарушением кроветворения, т. е. анемией.

Слайд 21Витамины относятся к жизненно необходимым компонентам пищи. Они нужны животным организмам лишь в очень небольших количествах для синтеза коферментов и сигнальных веществ
При нормальном питании суточная потребность организма в витаминах удовлетворяется полностью. Недостаточное или неполноценное питание (например, несбалансированная диета у пожилых людей, недостаточное питание у алкоголиков, потребление полуфабрикатов) или нарушение процессов усвоения и использования витаминов могут быть причиной различных форм витаминной недостаточности, вплоть до авитаминоза. Важная роль в обеспечении организма рядом витаминов (К, B12, H) принадлежит микрофлоре пищеварительного тракта. Поэтому дефицит витаминов может возникать вследствие медикаментозного лечения с использованием антибиотиков. Только немногие из витаминов, такие, как A, D, Е, В12, могут накапливаться в организме. Поэтому витаминная недостаточность быстро влечет за собой болезни витаминодефицита, затрагивающие состояние кожи, клетки крови и нервную систему организма. Витаминная недостаточность излечивается посредством полноценного питания или с помощью витаминных препаратов. Явление гипервитаминоза касается лишь витаминов А и D. Избыточное количество большинства других витаминов быстро выводится из организма с мочой.

Слайд 22По растворимости витамины подразделяются на жирорастворимые и водорастворимые. В химическом отношении жирорастворимые витамины А, D, E и К относятся к изопреноидам
Слайд 23Хромопротеиды
- Гемоглобин, синтез гемоглобина. - Распад гемоглобина, образование пигментов желчи, кала и мочи. - Нарушение пигментного обмена
Слайд 24Главная функция эритроцитов — транспорт кислорода от легких в ткани и СО2 от тканей обратно в легкие. Высшие организмы нуждаются для этого в специальной транспортной системе, так как молекулярный кислород плохо растворим в воде: в 1 л плазмы крови растворимо только около 3,2 мл О2. Содержащийся в эритроцитах белок гемоглобин (Hb) способен связать в 70 раз больше — 220 мл О2/л. Содержание Hb в крови составляет 140-180 г/л у мужчин и 120-160 г/л у женщин, т. е. вдвое выше по сравнению с белками плазмы (50-80 г/л). Поэтому Hb вносит наибольший вклад в образование рН-буферной емкости крови.
 является тетрамером, состоящим из двух α- и двух β-субьединиц с молекулярными массами примерно 16 кДа. α- и β-цепи отличаются аминокислотной последовательностью, но имеют сходную конформацию. Примерно 80% аминокислотных остатков гл")
Слайд 25Структура гемоглобина Гемоглобин взрослого организма (HbA, см. ниже) является тетрамером, состоящим из двух α- и двух β-субьединиц с молекулярными массами примерно 16 кДа. α- и β-цепи отличаются аминокислотной последовательностью, но имеют сходную конформацию. Примерно 80% аминокислотных остатков глобина образуют α-спирали, обозначенные буквами А-Н (см. схему). Каждая субъединица несет группу гема с ионом двухвалентного железа в центре. При связывании O2 с атомом железа в геме (оксигенация Hb) и отщеплении O2 (дезоксигенация) степень окисления атома железа не меняется. Окисление Fe2+ до Fe3+ в геме носит случайный характер. Окисленная форма гемоглобина, метгемоглобин, не способна переносить O2. Доля метгемоглобина поддерживается ферментами на низком уровне и составляет поэтому обычно только 1-2%. Четыре из шести координационных связей атома железа в гемоглобине заняты атомами азота пиррольных колец, пятая — остатком гистидина глобина (проксимальный остаток гистидина), а шестая — молекулой кислорода в оксигемоглобине и, соответственно, Н2О в дезоксигемоглобине.
: обозначаемых как Т- и R-формы соответственно. Т-Форма (напряженная от англ. tense) обладает существенно более низким сродством к O2 по сравнению с R-формой (на схеме с")
Слайд 26Аллостерические эффекты в гемоглобине Аналогично аспартат-карбамоилтрансферазе Hb может находиться в двух состояниях (конформациях): обозначаемых как Т- и R-формы соответственно. Т-Форма (напряженная от англ. tense) обладает существенно более низким сродством к O2 по сравнению с R-формой (на схеме справа). Связывание O2 с одной из субъединиц Т-формы приводит к локальным конформационным изменениям, которые ослабляют связь между субъединицами. С возрастанием парциального давления O2 увеличивается доля молекул Hb в высокоаффинной R-форме (от англ. relaxed). Благодаря кооперативным взаимодействиям между субъединицами с ростом концентрации кислорода повышается сродство Hb к O2, в результате чего кривая насыщения имеет сигмоидальный вид. На равновесие между Т- и R-формами влияют различные аллостерические эффекторы, регулирующие связывание O2 гемоглобином (желтые стрелки). К наиболее важным эффекторам относятся CO2, Н+ и 2,3-дифосфоглицерат [ДФГ (BPG)].
Слайд 27
Слайд 28Гем, железосодержащее тетрагидропиррольное красящее вещество, является составной частью О2-связывающих белков и различных коферментов оксидоредуктаз. Почти на 85% биосинтез гема происходит в костном мозге и лишь небольшая часть — в печени. В синтезе гема участвуют митохондрии и цитоплазма.
Слайд 29
, промежуточного продукта цитратного цикла, конденсацией с глицином получается продукт, декарбоксилирование которого приводит к 5-аминолевулинату (ALA). Отвечающая за эту стадию 5-аминолевулинат-синтаза (A")
Слайд 30Синтез тетрагидропиррольных колец начинается в митохондриях. Из сукцинил-КоА (на схеме наверху), промежуточного продукта цитратного цикла, конденсацией с глицином получается продукт, декарбоксилирование которого приводит к 5-аминолевулинату (ALA). Отвечающая за эту стадию 5-аминолевулинат-синтаза (ALA-синтаза) [1] является ключевым ферментом всего пути. Экспрессия синтеза ALA-синтазы тормозится гемом, т. е. конечным продуктом, и имеющимся ферментом. Это типичный случай торможения конечным продуктом, или ингибирования по типу обратной связи. После синтеза 5-аминолевулинат переходит из митохондрий в цитоплазму, где две молекулы конденсируются в порфобилиноген, который уже содержит пиррольное кольцо [2]. Порфобилиноген-синтаза [2] ингибируется ионами свинца. Поэтому при острых отравлениях свинцом в крови и моче обнаруживают повышенные концентрации 5-аминолевулината. На последующих стадиях образуется характерная для порфирина тетрапиррольная структура. Связывание четырех молекул порфобилиногена с отщеплением NH2-групп и образованием уропорфириногена III катализируется гидроксиметилбилан-синтазой [3]. Для образования этого промежуточного продукта необходим второй фермент, уропорфириноген III-синтаза [4]. Отсутствие этого фермента приводит к образованию «неправильного» изомера — уропорфириногена I. Тетрапиррольная структура уропорфиринoгена III все еще существенно отличается от гема. Так, отсутствует центральный атом железа, а кольцо содержит только 8 вместо 11 двойных связей. Кроме того, кольца несут только заряженные боковые цепи R (4 ацетатных и 4 пропионатных остатков). Так как группы гема в белках функционируют в неполярном окружении, необходимо, чтобы полярные боковые цепи превратились в менее полярные. Вначале четыре ацетатных остатка (R1) декарбоксилируются с образованием метильных групп (5). Образующийся копропорфириноген III снова возвращается в митохондрии. Дальнейшие стадии катализируются ферментами, которые локализованы на/или внутри митохондриальной мембраны. Прежде всего под действием оксидазы две пропионатные группы (R2) превращаются в винильные (6). Модификация боковых цепей заканчивается образованием протопорфириногена IX.

Слайд 31
![Деградация гемоглобина В организме человека в течение 1 ч разрушается примерно 100-200 млн эритроцитов. Разрушение начинается в микросомальной фракции ретикуло-эндотелиальной системы [РЭС (RES)] клеток печени, селезенки и костного мозга. После отделения белковой части (глобина) красный гем расщепляе](https://prezentacii.org/upload/cloud/19/02/115794/images/thumbs/screen32.jpg "Деградация гемоглобина В организме человека в течение 1 ч разрушается примерно 100-200 млн эритроцитов. Разрушение начинается в микросомальной фракции ретикуло-эндотелиальной системы [РЭС (RES)] клеток печени, селезенки и костного мозга. После отделения белковой части (глобина) красный гем расщепляе")
Слайд 32Деградация гемоглобина В организме человека в течение 1 ч разрушается примерно 100-200 млн эритроцитов. Разрушение начинается в микросомальной фракции ретикуло-эндотелиальной системы [РЭС (RES)] клеток печени, селезенки и костного мозга. После отделения белковой части (глобина) красный гем расщепляется гем-оксигеназой с помощью кислорода и НАДФН на ионы Fe2+, СО (оксид углерода!) и зеленый биливердин. Далее железо утилизируется. Затем биливердин восстанавливается биливердинредуктазой до оранжевого билирубина. Это изменение цвета легко можно наблюдать in vivo в виде синяков (гематомах). Интенсивный цвет гема и других порфиринов является результатом сопряжения многочисленных двойных связей, которые образуют две резонансно стабилизированные (мезомерные) системы. Для дальнейшего разрушения билирубин транспортируется кровью в печень. Так как он плохо растворим в плазме, транспорт осуществляется в комплексе с альбумином. В том же участке связывания альбумина сорбируются и лекарственные препараты. Паренхиматозные клетки печени забирают билирубин из крови. После того как билирубин в печени дважды конъюгируется с активированной глюкуроновой кислотой (УДФ-GIcUA (не показано), повышается его водорастворимость. Образование конъюгата катализируется УДФ-глюкуронозилтрансферазой — ферментом, находящимся в ЭР печени, а также в незначительных количествах в почках и слизистой кишечника. Глюкуроновая кислота присоединяется к пропионатным боковым цепям билирубина сложноэфирными связями. Образующийся диглюкуронид билирубина переносится в желчь путем активного транспорта против градиента концентрации. Этот транспорт является скорость-лимитирующей стадией метаболической трансформации билирубина в печени. Лекарственные препараты, такие, как, например фенобарбитал, могут индуцировать образование конъюгата и транспортный процесс. В кишечнике конъюгат билирубина снова частично расщепляется бактериальной β-глюкуронидазой. Свободный билирубин постепенно восстанавливается до бесцветного уробилиногена и стеркобилиногена, которые далее окисляются кислородом воздуха до уробилина и стеркобилина. Эти конечные продукты метаболической трансформации желчных пигментов в кишечнике окрашены в цвета от оранжевого до желтого. Они выделяются по большей части с калом, а в меньшей степени резорбируются (энтерогепатическая циркуляция). При интенсивном процессе разрушения гема в моче внезапно появляется уробилиноген, где он при окислении кислородом воздуха темнеет, превращаясь в уробилин. Наряду с гемоглобином, по аналогичному пути разрушаются группы гема и у других гемсодержащих белков (миоглобина, цитохрома, каталазы, пероксидазы). Однако их вклад в образование желчных пигментов (250 мг в сутки) составляет лишь примерно 10-15%.

Слайд 34Нарушение пигментного обмена
Желтуха представляет собой желтое неестественное окрашивание кожи или склер. Это связано с присутствием в плазме билирубина в концентрациях, превышающих 40 мкмоль/л. В норме концентрация билирубина в плазме менее 22 мкмоль/л.
Слайд 35Имеются три основные причины повышения уровня билирубина в крови: - Скорость синтеза билирубина повышена и превышает выделительную способность печени (гемолитическая, надпеченочная желтуха). - Угнетение конъюгационных и/или выделительных механизмов в печени — снижается способность печени метаболизировать синтезируемый в нормальных количествах билирубин (печеночная, гепатоцеллюлярная желтуха). - Обструкция билиарной системы, препятствующая оттоку желчи (холестатическая, подпеченочная, механическая, обструкционная желтуха).
Слайд 36Нарушение пигментного обмена. Причины желтухи
Слайд 37
Слайд 38Нарушение пигментного обмена. Дифференциальная диагностика желтух
Слайд 39
 Слайд 1
Слайд 1 Слайд 2
Слайд 2 Слайд 3
Слайд 3 Слайд 4
Слайд 4 Слайд 5
Слайд 5 Слайд 6
Слайд 6 Слайд 7
Слайд 7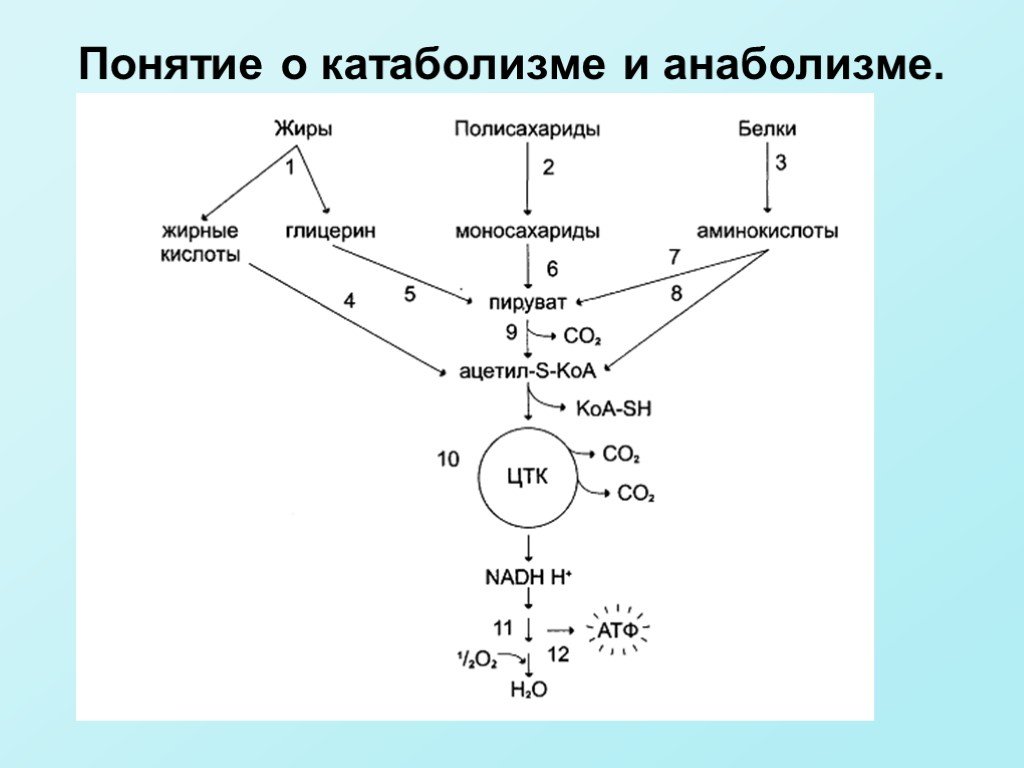 Слайд 8
Слайд 8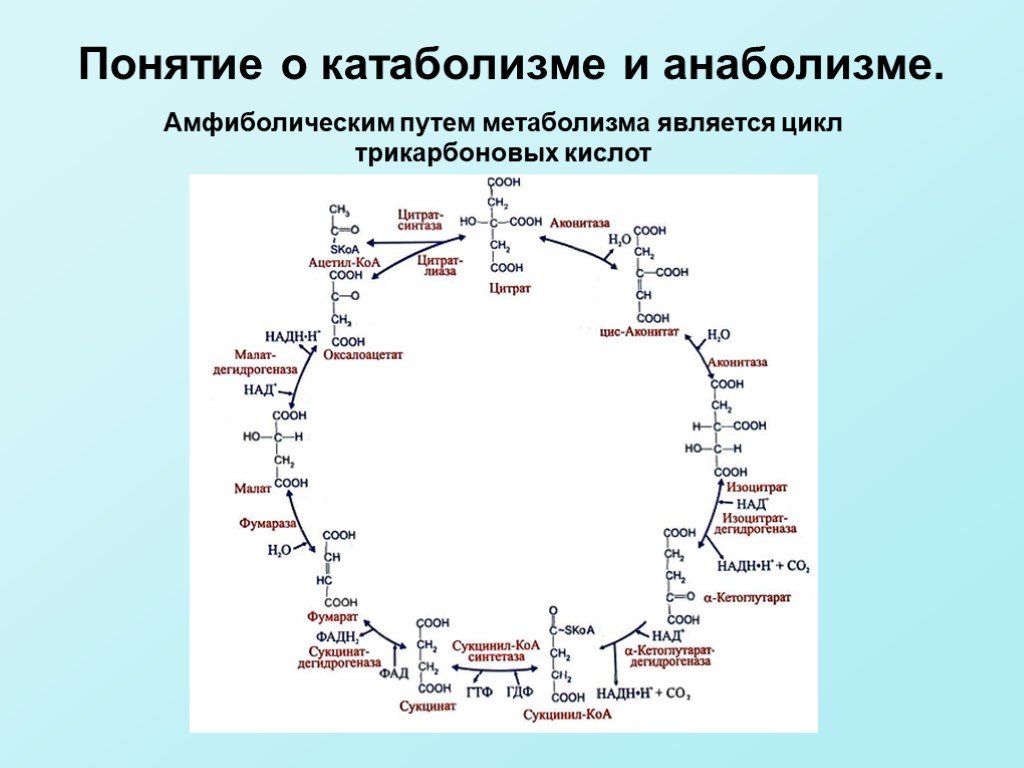 Слайд 9
Слайд 9 Слайд 10
Слайд 10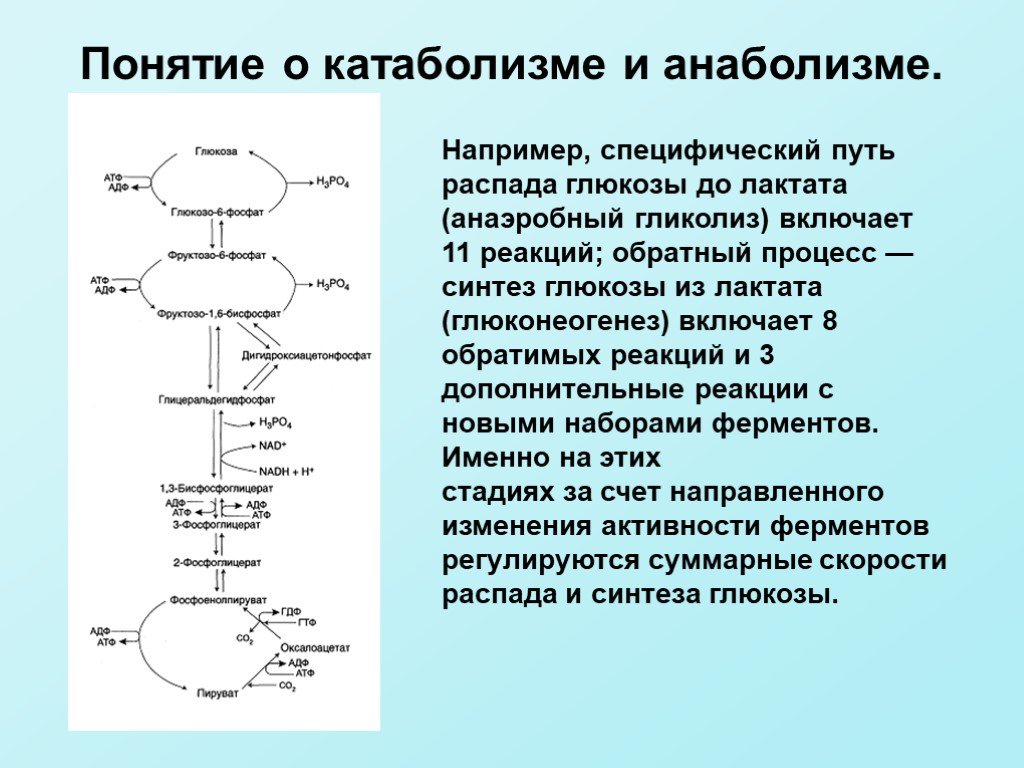 Слайд 11
Слайд 11 Слайд 12
Слайд 12 Слайд 13
Слайд 13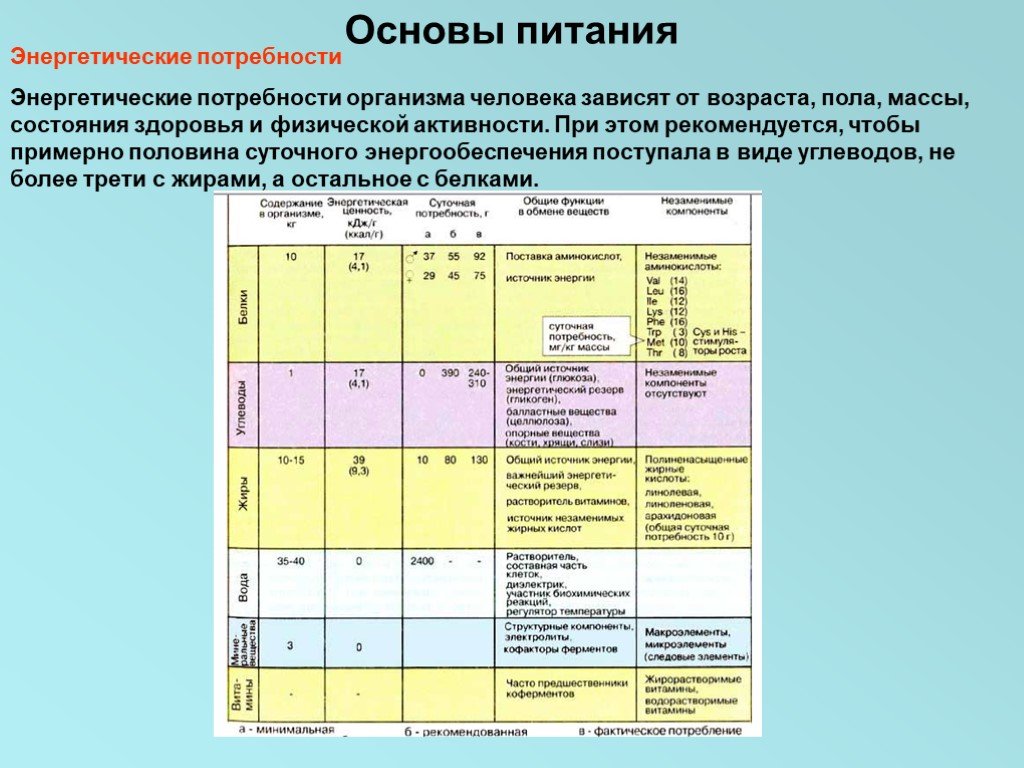 Слайд 14
Слайд 14 Слайд 15
Слайд 15 Слайд 16
Слайд 16 Слайд 17
Слайд 17 Слайд 18
Слайд 18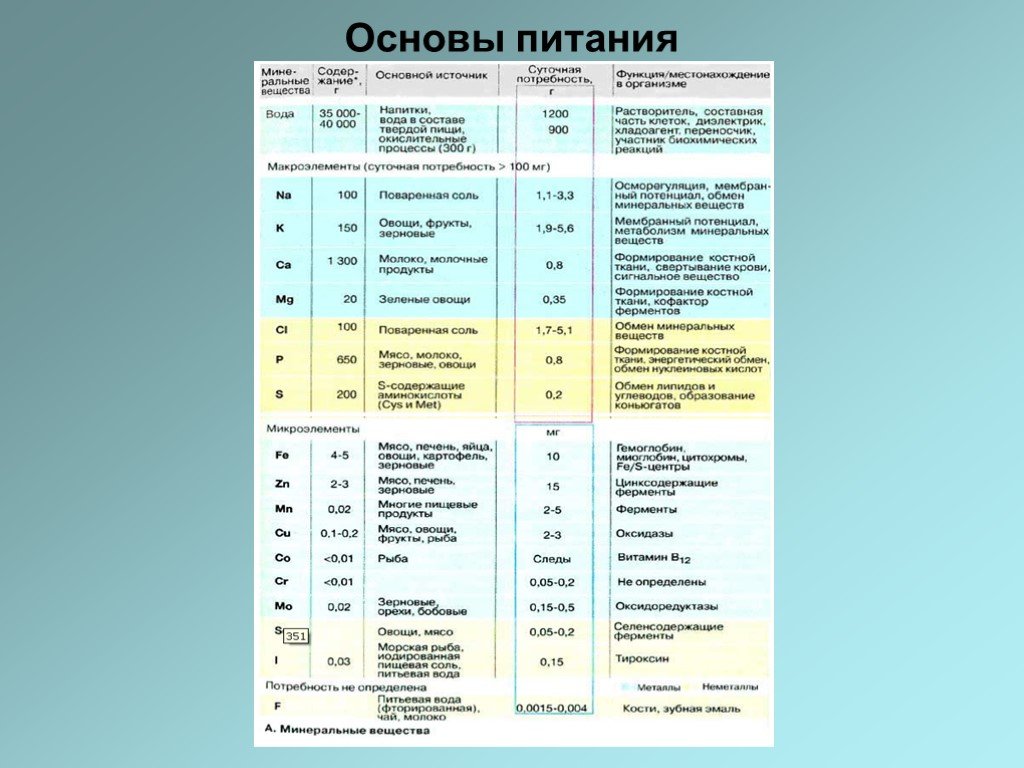 Слайд 19
Слайд 19 Слайд 20
Слайд 20 Слайд 21
Слайд 21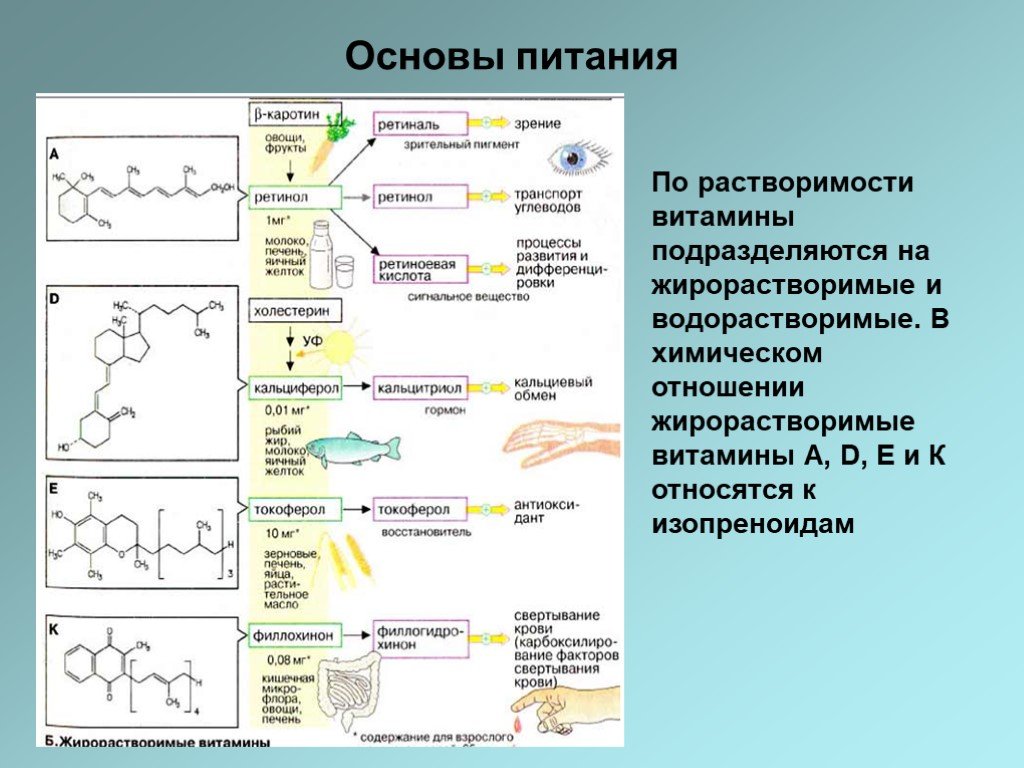 Слайд 22
Слайд 22 Слайд 23
Слайд 23 Слайд 24
Слайд 24 Слайд 25
Слайд 25 Слайд 26
Слайд 26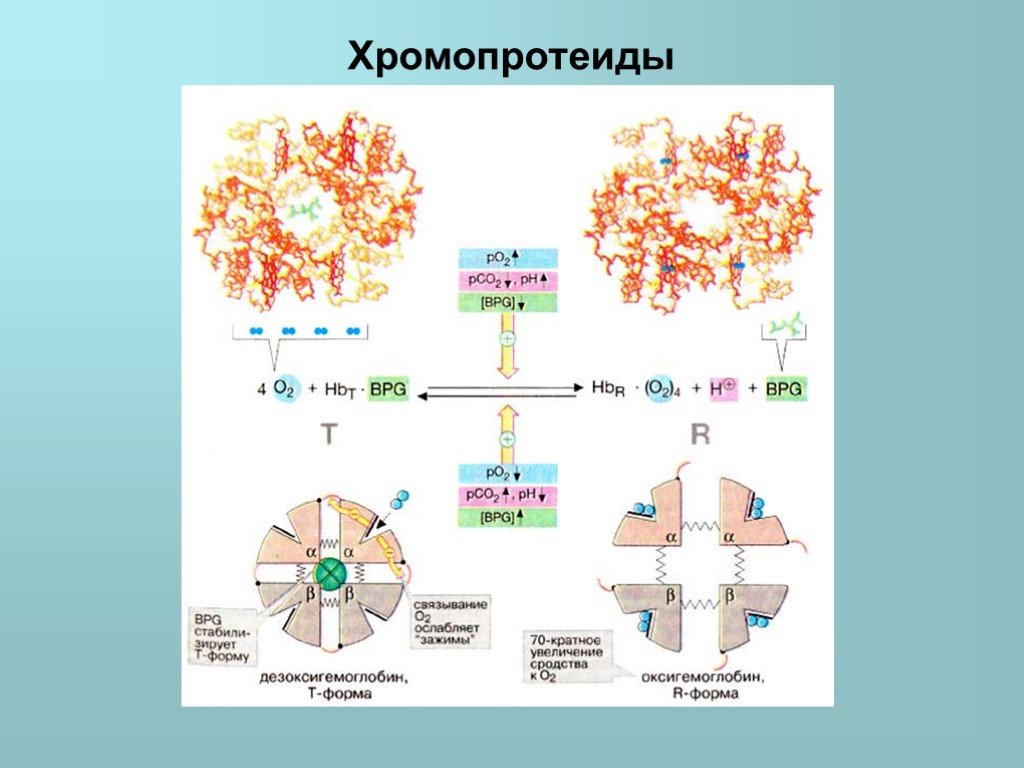 Слайд 27
Слайд 27 Слайд 28
Слайд 28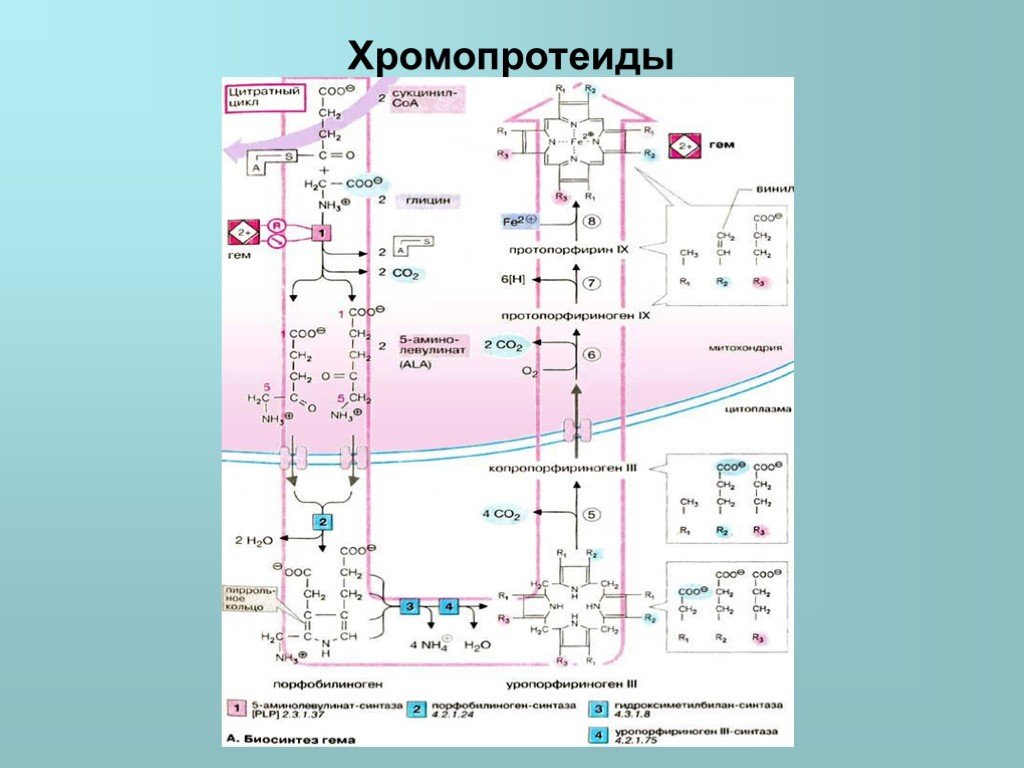 Слайд 29
Слайд 29 Слайд 30
Слайд 30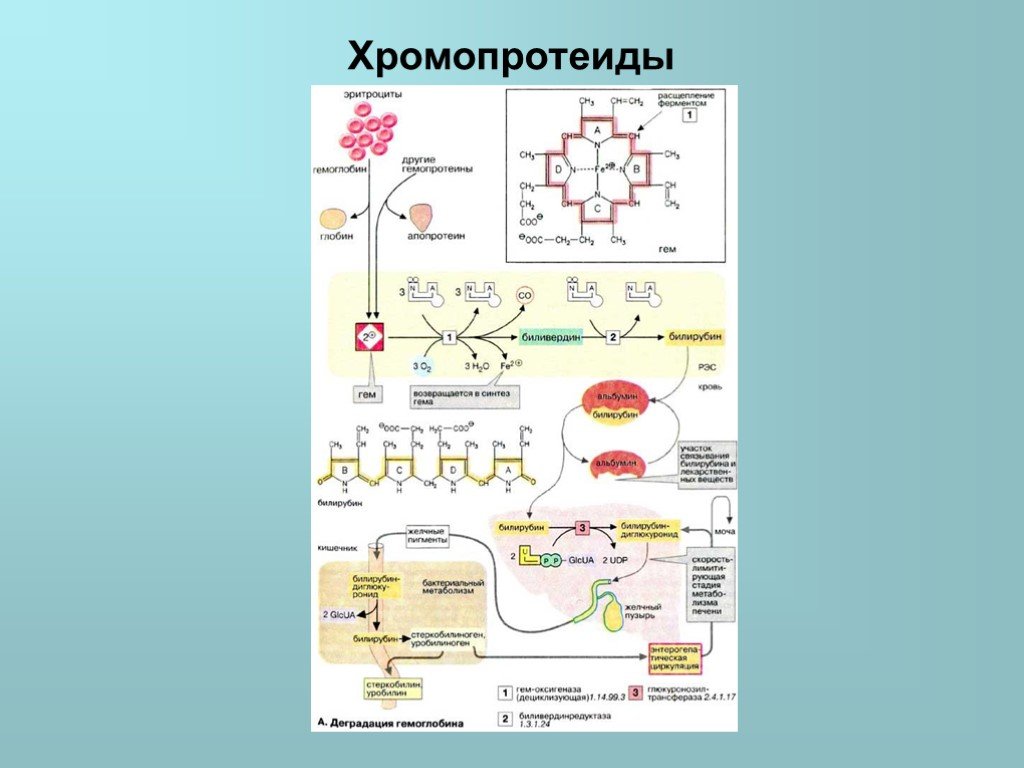 Слайд 31
Слайд 31 Слайд 32
Слайд 32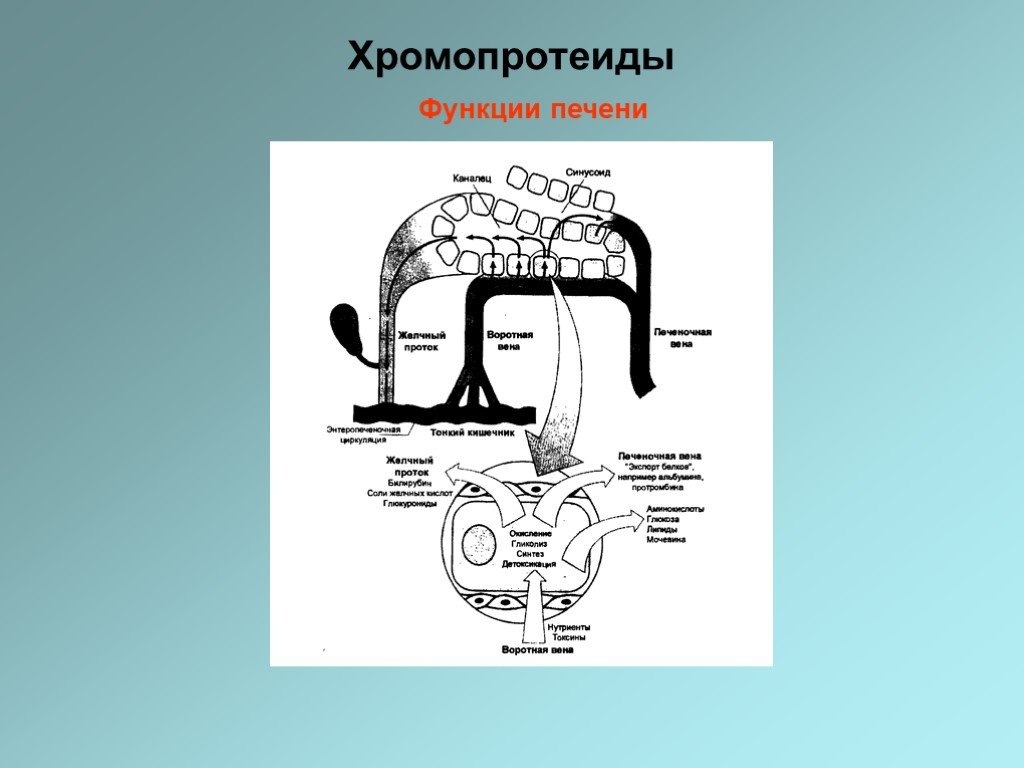 Слайд 33
Слайд 33 Слайд 34
Слайд 34 Слайд 35
Слайд 35 Слайд 36
Слайд 36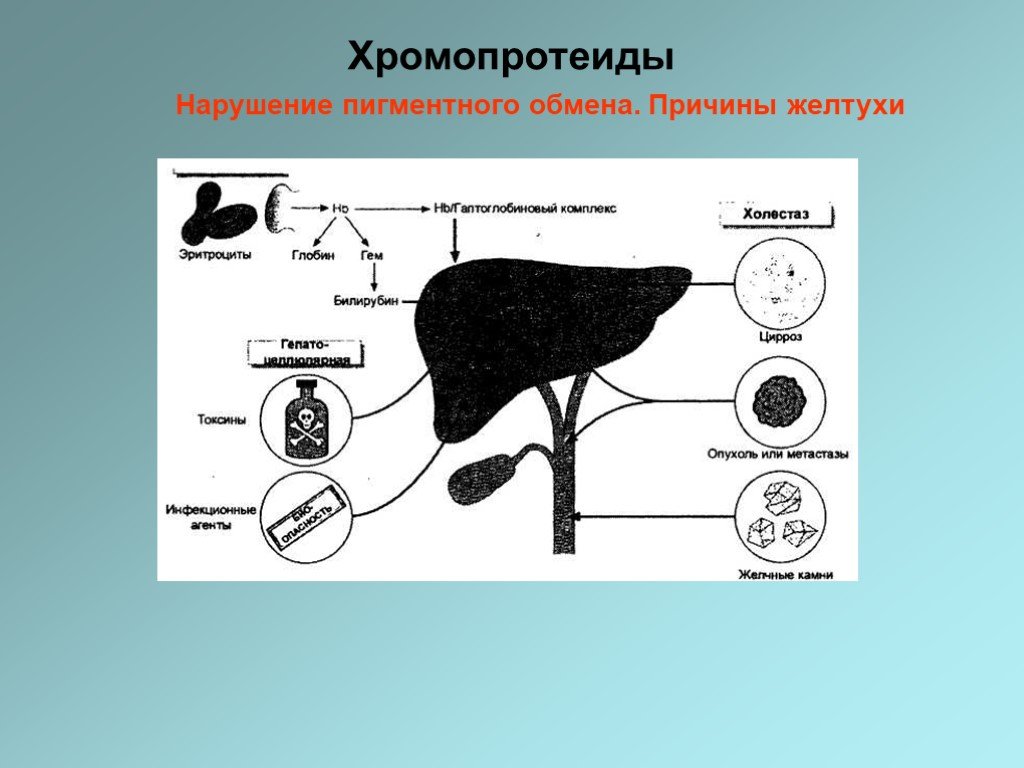 Слайд 37
Слайд 37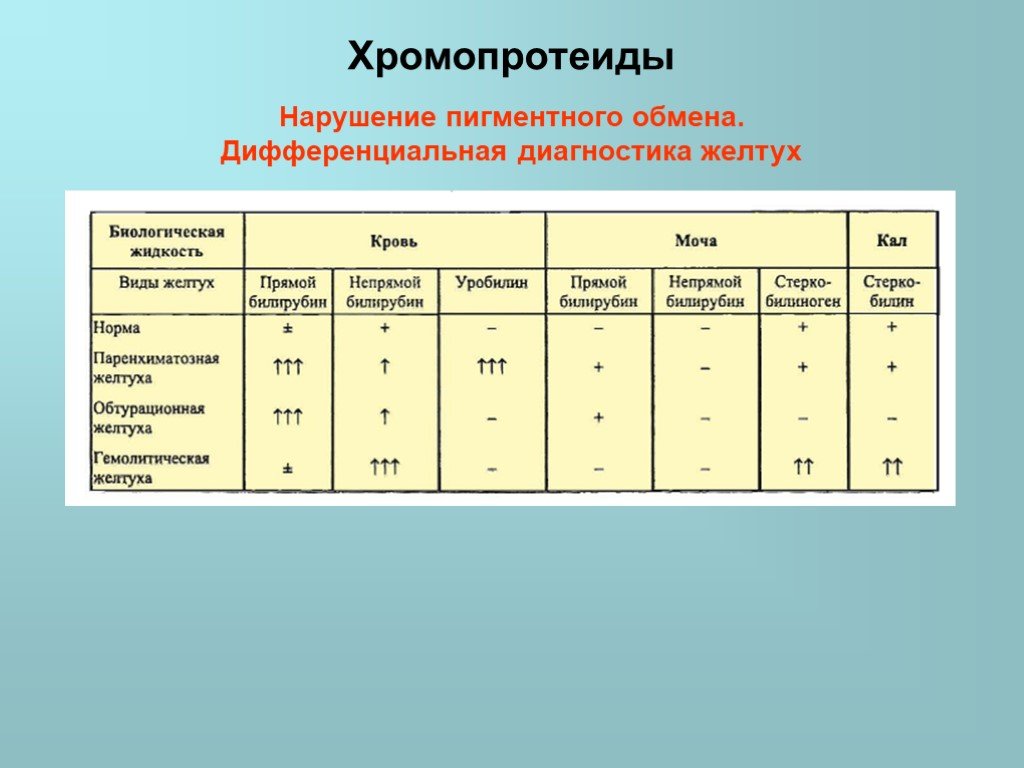 Слайд 38
Слайд 38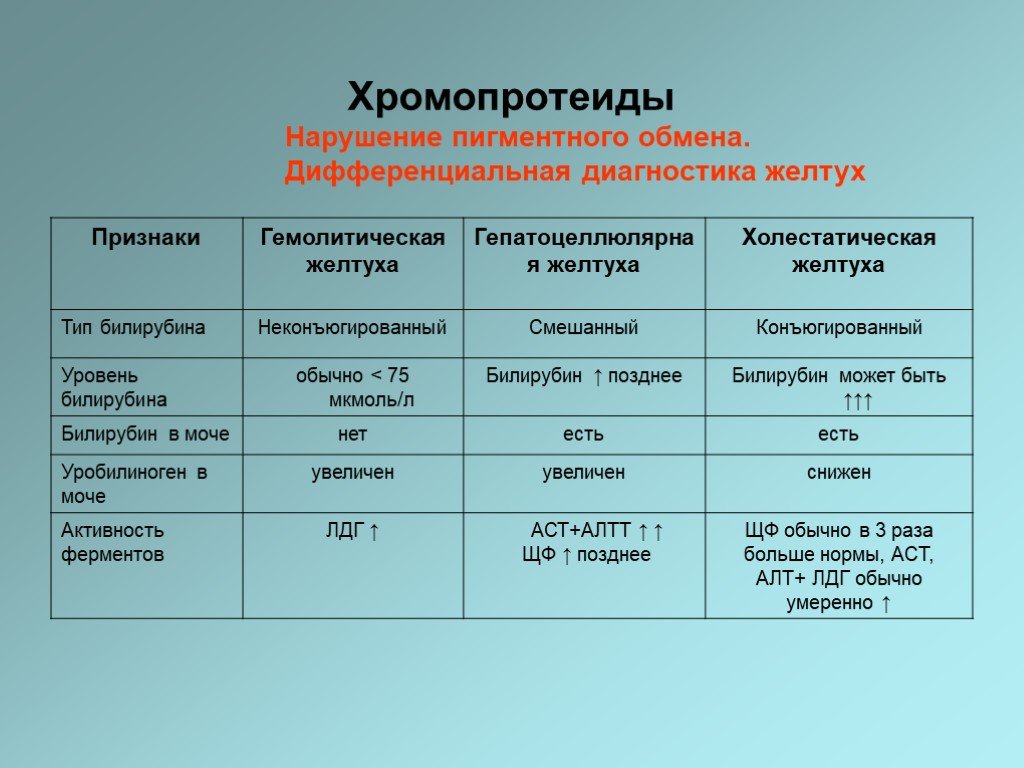 Слайд 39
Слайд 39




















")







