Слайд 1Противобактериальный иммунный ответ
Сидорова А. А.
. Бактерии и их токсины нейтрализуются антибактериальными и антитоксическими антителами. Комплексы бактерия (антигены)-антитела активируют комплемент, компоненты которого присоединяются")
Слайд 2Противобактериальный иммунитет направлен как против бактерий, так и против их токсинов (антитоксический иммунитет). Бактерии и их токсины нейтрализуются антибактериальными и антитоксическими антителами. Комплексы бактерия (антигены)-антитела активируют комплемент, компоненты которого присоединяются к Fc-фрагменту антитела, а затем образуют мембраноатакующий комплекс, разрушающий наружную мембрану клеточной стенки грамотрицательных бактерий. Пептидогликан клеточных стенок бактерий разрушается лизоцимом. Антитела (Fc-фрагмент) и комплемент (С3b), обволакивают бактерии и "приклеивают" их к Fc- и С3b-рецепторам фагоцитов, выполняя роль опсонинов вместе с другими белками, усиливающими фагоцитоз (С-реактивным белком, фибриногеном, маннансвязывающим лектином, сывороточным амилоидом). Фагоцитоз является основным механизмом антибактериального иммунитета. Фагоциты направленно перемещаются к объекту фагоцитоза, реагируя на хемоаттрактанты вещества микробов, активированные компоненты комплемента (С5а, СЗа) и цитокины. Противобактериальная защита слизистых оболочек обусловлена секреторными IgA, которые, взаимодействуя с бактериями, препятствуют их адгезии на эпителиоцитах.

Слайд 3Локализация патогенов и эффективные пути иммунной защиты

Слайд 4Поверхностные структуры микроорганизмов и их взаимодействие с рецепторами клеток иммунной системы
Антигенная структура бактерий определяется главным образом осо- бенностями строения их клеточной стенки. Основу клеточной стенки всех микроорганизмов составляют пептидогликаны, в состав которых входят остатки мурамиловой кислоты и пептидные компоненты. У грамположи- тельных бактерий мурамилпептиды являются поверхностными структура- ми. На поверхности грамотрицательных бактерий поверх мурамилпептидов локализуется липидный бислой, в составе которого присутствуют ЛПС, играющие роль эндотоксинов. Основа ЛПС — полисахарид, соединенный с короткими группами липида А, обусловливающего токсичность ЛПС. С ним связано также большинство проявлений биологической активнос- ти ЛПС. Поверхность микобактерий имеет дополнительные компоненты в виде гликолипидов, миколевой кислоты, липоарабиноманнана и т.д. Своими особенностями отличаются структуры поверхностей грибков, про- стейших, риккетсий и других микроорганизмов, выступающих в качестве патогенов.
. Фагоцитоз и разрушение ба")
Слайд 7Защита от внеклеточных бактерий
Бактерии распознаются TLR макрофагов, тучных, эпителиальных и других клеток. Активация клеток, секреция провоспалительных цитокинов и формирование воспалительной реакции, сопровождающейся миграцией из сосудов в очаг лейкоцитов (нейтрофилов). Фагоцитоз и разрушение бактерий. При ограниченном количестве патогена иммунная защита успешно реализуется с помощью реакций врожденного иммунитета. В первую линию защиты вовлекаются NKT-клетки (секретируют IFNy), убТ-клетки (участвуют в бактериолизе с помощью невыясненных механизмов) Важнейший фактор ранней защиты против внеклеточных патогенов - естественные антитела пресинтезированные В1 -клетками.
 специфичные к распространенным АГ бактерий –фосфорилхолину,липополисохариду,пептидогликану и др. Связывание АТ с бактериями –активация С* по классическому пути, опсонизация бактерий (их фагоцитоз), а иногда (нейссерии)-лизис бактерий. Внеклеточные микроорганизмы - действие фактор")
Слайд 8Нормальные АТ (IgM) специфичные к распространенным АГ бактерий –фосфорилхолину,липополисохариду,пептидогликану и др. Связывание АТ с бактериями –активация С* по классическому пути, опсонизация бактерий (их фагоцитоз), а иногда (нейссерии)-лизис бактерий. Внеклеточные микроорганизмы - действие факторов врожденного иммунитета: -пентраксины (опсонизия бактерий и активация С*), дефензинц секретируемые эпителиальными клеткамии фагоцитами. Активация комплемента на их поверхности происходит не только по классическому, но и по альтернативному пути. Резерв врожденного иммунитета достаточен для отражения большинства атак внеклеточных бактерий.
 и доставляют в региональный лимфоузел Презентация АГ-пептида СD4+ Т-лимфоцитам Активация Th2-клеток В очагах поражения и лимфатических узлах В-лимфоциты распознают АГ бактерий, обрабатывают и презентиру")
Слайд 9Адаптивный антибактериальный иммунитет
ДК захватывают бактерии и их продукты (экзотоксины и др.) и доставляют в региональный лимфоузел Презентация АГ-пептида СD4+ Т-лимфоцитам Активация Th2-клеток В очагах поражения и лимфатических узлах В-лимфоциты распознают АГ бактерий, обрабатывают и презентируют специфическим Th2-клетками. В-клетки получают от Т-хелперов сигнал через костииулируюцую молекулу CD40. IL-4, секретируемый ТН2, обепечивает пролиферацию клона активированных В-лимфоцитов. В лимфоидных фолликулах при участии фолликулярных ДК происходит формирование зародышевых центров. В них мигрирует большинство стимулированных В-лимфоцитов, а также фолликулярные CXCR5+ Т-хелперы.
 клеток. Миграция ПК в кр")
Слайд 10Пролиферация и переключение изотипов BCR и повышение его сродства к АГ в В-клетках и отбор клонов по сродству к АГ, представленному в составе иммунных комплексов на фолликулярных ДК. Миграция В-клеток в апикальную зону зародышевых центров дифференцировка плазматических (АОК) клеток. Миграция ПК в красную пульпу селезенки, мозговые шнуры лимфоузлов и в костный мозг, где они секретируют АТ. AT (IgA) секретируются в мукозальном отделе ИС
Слайд 11АТ взаимодействует с АГ на поверхности. внеклеточных патогенов. АТ против жгутиковых антигенов, обездвиживают клетку. IgА-антитела, связывающиеся с бактериями в просвете кишечника, препятствуют их проникновению через кишечную стенку. Взаимодействуя с токсинами, АТ вызывают их инактивацию (нейтрализация). Защитный эффект антител реализуется также с участием фагоцитов-макрофагов (эффект опоснизации) или С’ (эффекты оспонизации и лизиса).
, разная степень подавления у них клеточного иммунитета. Факторы первой линии в защите от микобактерий: участие в ранних эффекторных реакциях")
Слайд 12Адаптивный иммунитет к внутриклеточным бактериям
Иммунная защита против Мicobscteriwn tuberculosis. Развитие заболевания происходит не у всех больных (25-40%), разная степень подавления у них клеточного иммунитета. Факторы первой линии в защите от микобактерий: участие в ранних эффекторных реакциях убТ- лимфоцитов. Микобактерии взаимодействуют с альвеолярными макрофагами. Гпиколипиды и липотейхоевая кислота клеточной стенки воздействуют на рецептор TLR-2, а ЛПС- на TLR-4. Фактор вирулентности микобактерий липоарабиноманнан (LAM) взаимодействует с концевыми остатками маннозы с формированием комплекса LAM-Man, который посредством фосфатаз подавляет внутриклеточную передачу сигнала от TLR.

Слайд 13Защитные механизмы бактерий
1. Подавление хемотаксиса 2.Капсула или оболочка препятствуют связыванию бактерии фагоцитами. 3.Блокада запуска бактерицидных механизмов, М. tuberculosis ингибирует слияние лизосом с фагоцитами, а также подкисляющий содержимое фагосом протонный насос и в результате pH в них не снижается. 4.Каталаза разрушает образуемый фагоцитами пероксид водорода. 5.Клетки М.lrprae окружены фенольным гликолипидом, который инактивирует свободные радикалы. 6. Микобактерии выделяют липоарабиноманнан, блокирующий активацию макрофагов на Ифу 7. Утрата функции презентации АГ инфицированным фагоцитами. 8. Бактерии выходят из фагосом и размножаются в цитоплазме фагоцита , который погибает.
Слайд 14Особенности иммунитета при бактериальных инфекциях
Аллергизация по типу UPN Ат-зависимая цитотоксичность (активированные макрофаги и NK-клетки)
Слайд 15Спасибо за внимание ._.
 Слайд 1
Слайд 1 Слайд 2
Слайд 2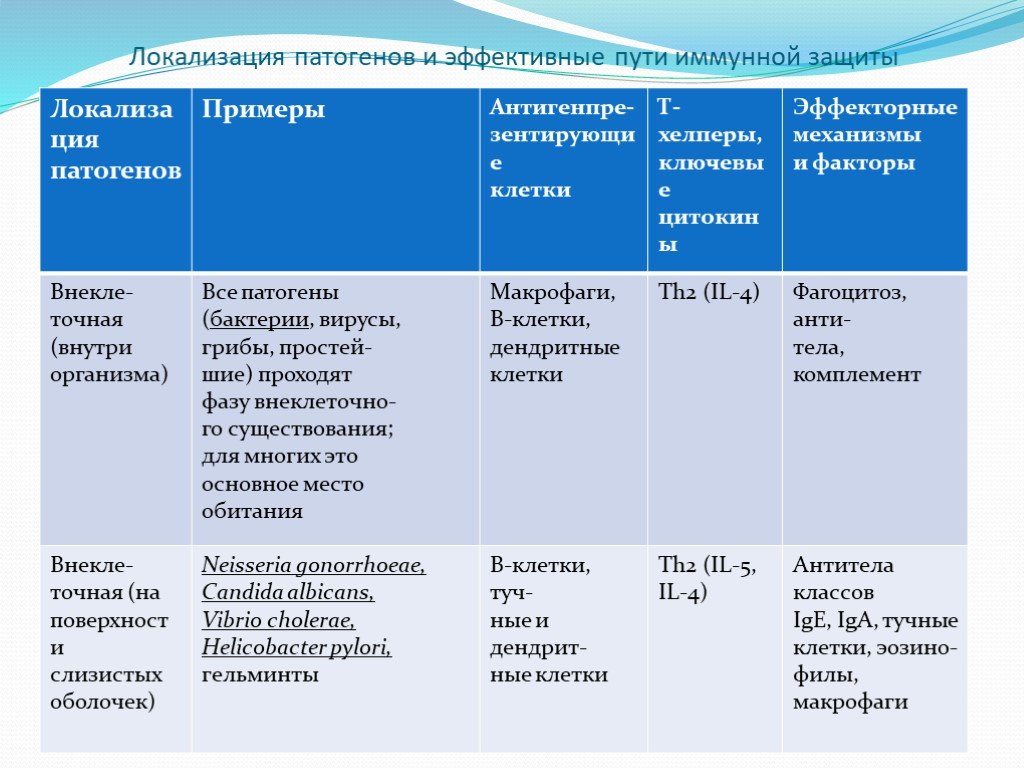 Слайд 3
Слайд 3 Слайд 4
Слайд 4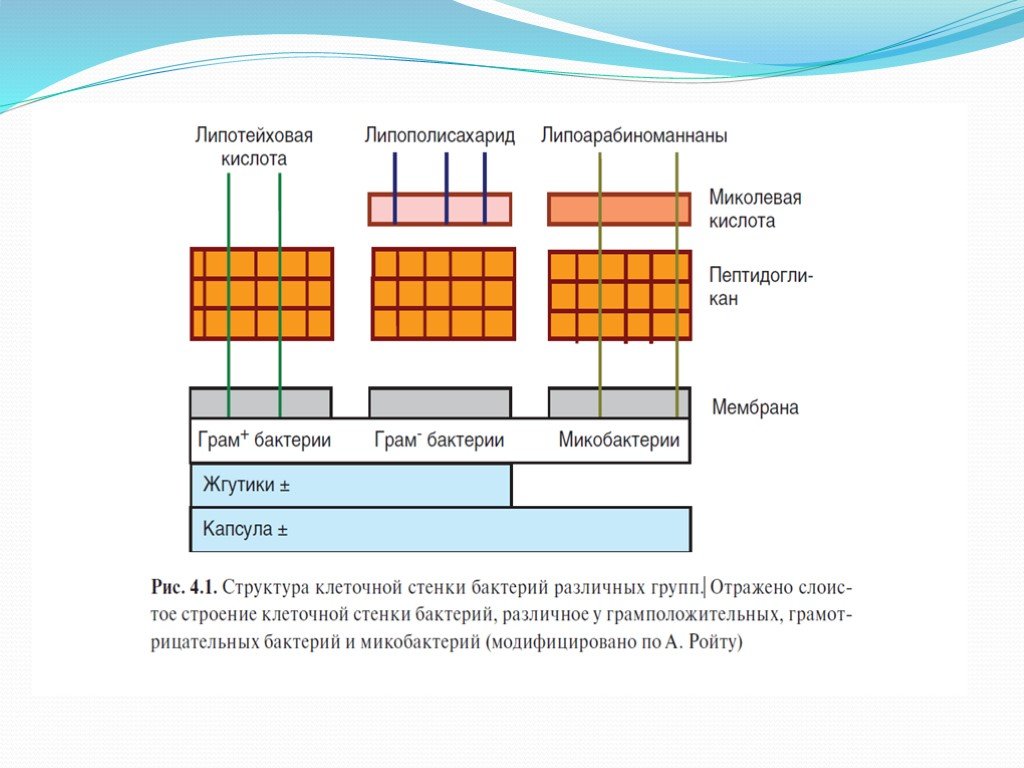 Слайд 5
Слайд 5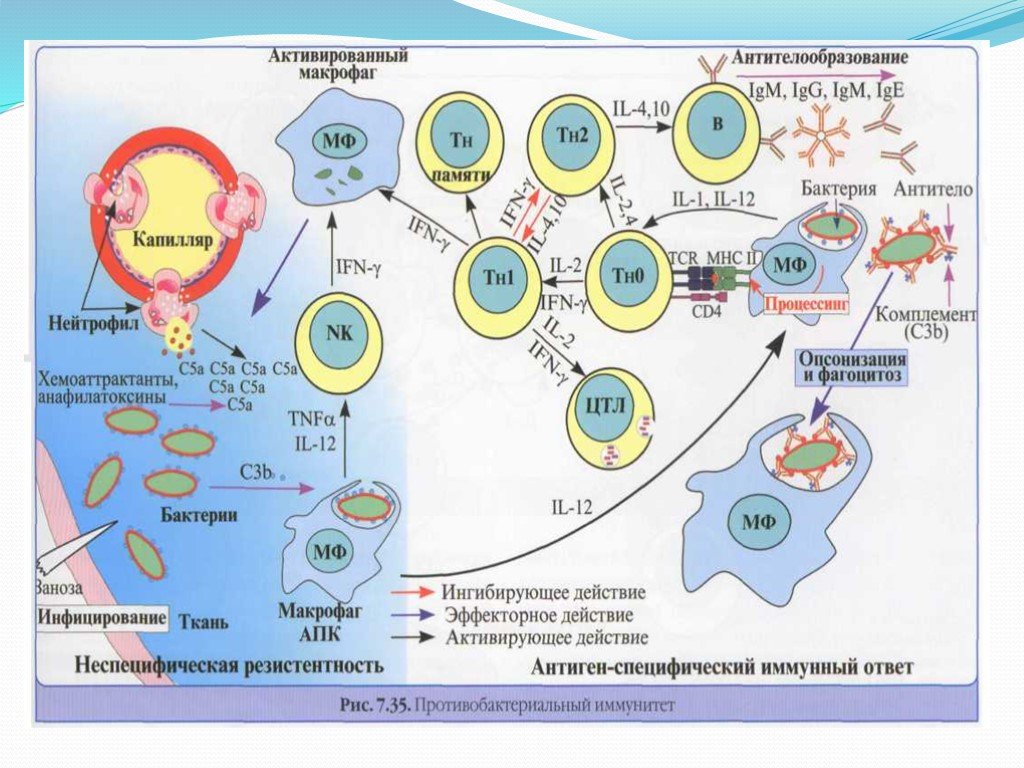 Слайд 6
Слайд 6 Слайд 7
Слайд 7 Слайд 8
Слайд 8 Слайд 9
Слайд 9 Слайд 10
Слайд 10 Слайд 11
Слайд 11 Слайд 12
Слайд 12 Слайд 13
Слайд 13 Слайд 14
Слайд 14 Слайд 15
Слайд 15










